Polypodiidae
| Polypodiidae | ||
|---|---|---|
| Rango temporal: Pérmico-reciente | ||
 | ||
 Órganos y ciclo de vida de un helecho típico. | ||
| Taxonomía | ||
| Reino: | Plantae | |
| División: | Monilophyta | |
| Clase: | Filicopsida | |
| Subclase: | Polypodiidae | |
| Órdenes | ||
Según Christenhusz et al. 2011[1][2][3] (basada en Smith et al. 2006,[4] 2008);[5] que también provee una secuencia lineal de las licofitas y monilofitas. | ||
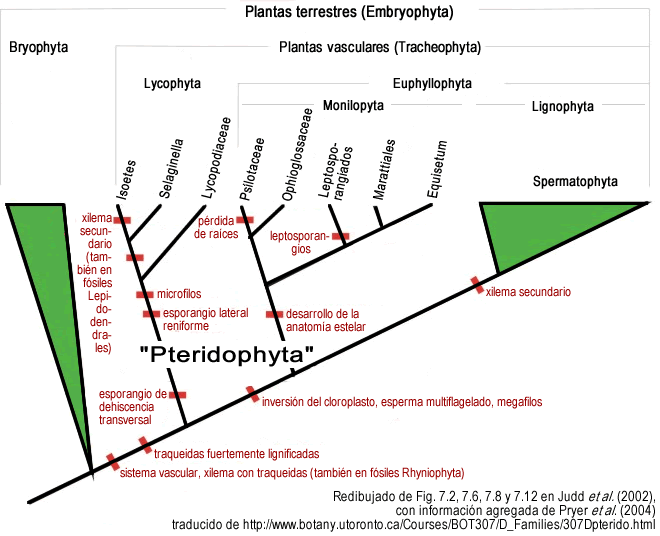
- Ver Pteridophyta para una introducción a las plantas vasculares sin semilla
Los helechos leptosporangiados (subclase Polypodiidae o clase Polypodiopsida, según el sistema de clasificación) se caracterizan por poseer un esporangio que se desarrolla a partir de una única célula, y en el corte del esporangio maduro se observa que su pared es de una célula de espesor. El esporangio nace en un "pie" que es una o más filas de células que lo separan del megafilo, y una parte de la pared se diferencia en un característico "anillo", que es una fila de células de la pared diferenciadas, que poseen las paredes internas engrosadas y la pared externa adelgazada. Cuando el anillo del esporangio se seca, las células del anillo se retraen por la parte externa, ejerciendo presión sobre el resto de la pared del esporangio y forzando su apertura. A este tipo de esporangio, característico y distintivo de este clado, se lo llama leptosporangio, y la morfología del anillo nos puede decir mucho acerca de las relaciones filogenéticas dentro del clado.
Dentro de polypodiopsida, se considera más antiguo al "anillo" de Osmundales (a veces llamado escudo de dehiscencia), le siguen los anillos "enteros", no interrumpidos en todo su recorrido, de los órdenes siguientes, y finalmente es más reciente (y más eficiente para la dehiscencia) el anillo de Polypodiales, que es el clado más grande (en el que se encuentran más de 12.000 especies vivientes), que es un anillo incompleto, interrumpido por una estructura especializada en la dehiscencia llamada estomio.
En el caso particular de los helechos acuáticos heterospóricos ("Salviniales" sensu Smith et al. 2006), que es un clado con muchas modificaciones sufridas probablemente debido a la presión de selección para la vida acuática, el anillo fue perdido en forma secundaria (no se observa, o se observa solo en forma relictual, pero sus antepasados lo poseían).
Los leptosporangios de la mayoría de las especies contienen un número relativamente pequeño y definido de esporas, en comparación con la cantidad producida por los eusporangios de Marattiopsida, Equisetopsida y Psilotopsida.
Los órdenes aquí presentados fueron altamente consensuados por los análisis moleculares recientes, por lo que probablemente ésta sea una clasificación definitiva de los mismos. Se desprenden en forma gradual del lineaje que deriva en los polypodiales.
Los Polypodiales son el orden más diverso de las polypodiopsidas, y los más recientes también, el registro fósil de los polypodiales comienza en el Terciario, y probablemente se diversificaron en forma oportunista explotando los nuevos nichos ecológicos creados por el establecimiento de las angiospermas.
Filogenia
[editar]- Introducción teórica en Filogenia
Los diferentes órdenes de los helechos leptosporangiados se relacionan filogenéticamente del siguiente modo:[6]
| Polypodiidae |
| ||||||||||||||||||||||||||||||||||||
Como circunscripto según Christenhusz et al. 2011[1][2][3] (basada en Smith et al. 2006,[4] 2008);[5] que también provee una secuencia lineal de las licofitas y monilofitas, (ver taxobox), el grupo es monofilético (Pryer et al. 2001). Según estos estudios, las maratiáceas son hermanas de las equisetáceas, pero con parentescos exactos todavía no dilucidados a ese momento, requiriendo consenso. Estas 3 clases son a su vez hermanas de Ophioglossaceae + Psilotaceae.
El árbol filogenético que muestra su relación entre ellos y con los clados hermanos de monilofitas ha sido el siguiente (véase también Pterophyta):
 Árbol filogenético y taxonomía de las clases y órdenes de monilophytas. Dibujado a partir de Smith et al. (2006). |
Diversidad de polypodiopsidas
[editar]Esta es una lista de los representantes más definidos de cada grupo de polypodiopsidas, creada con el fin de mostrar la diversidad necesaria para comprender los avances en taxonomía y filogenia de polypodiopsidas. Las descripciones de cada grupo son deliberadamente incompletas. Para una lista completa de taxones, o una explicación más detallada de los mismos, siga el árbol taxonómico en la ficha presentada al principio de esta página, o ingrese al artículo principal indicado en cada artículo.
Todas las polypodiopsidas son helechos con megafilos (frondes) de prefoliación circinada, y esporangio de tipo leptosporangiado (con pie, con anillo, con pared de 1 célula de espesor, con pocas esporas por esporangio). Las excepciones se encuentran en los clados adaptados a condiciones atípicas, como las que encontramos en los helechos acuáticos.
Osmunda y afines
[editar]|
Las osmundáceas son fácilmente reconocibles porque sus frondes tienen algunas pinas fértiles (más pequeñas y no fotosintéticas) y algunas pinas estériles fotosintéticas. ("trofoesporofilos sectoriales") Las pinas fértiles se encuentran en la parte apical o en la parte media de la fronde. También tienen estípulas (evaginaciones del tallo alrededor del pecíolo de las hojas) que ayudan a su determinación. Carácter de importancia taxonómica: Los de las osmundáceas son leptosporangios únicos, no típicos: si bien son leptosporangios, son considerados "primitivos", con pie de muchas células de espesor, con escudo de dehiscencia (parche de la pared del esporangio con células de pared engrosada) que ahora prefiere llamarse "anillo lateral" por su homología con el anillo de las demás polypodiopsidas.
|


Hymenophyllum y afines
[editar]|
Estos helechos son fácilmente reconocibles debido a que sus frondes poseen una lámina monoestratificada (de una sola capa de células de espesor) que les da un aspecto semitransparente. Son epífitos (usan otras plantas más altas como sustrato, sin parasitarlas) o saxícolas (usan las piedras como sustrato), se los encuentra en climas desde tropicales a templado-fríos, pero siempre en bosques con precipitaciones permanentes o selvas nubladas. Absorben agua no solo por las raíces sino por toda la superficie de su cuerpo. En español también se los conoce como "helechos reviviscentes", porque ante una reducción de la humedad su lámina se deseca, encogiéndose y recurvándose a modo de resistencia, de esta forma resisten la sequía, recuperando su forma cuando la humedad reaparece. Carácter de importancia taxonómica: El anillo del leptosporangio de las hymenofiláceas es transversal-oblicuo. Este tipo de anillo se considera más primitivo que los anillos verticales que veremos después. |
 |
Gleichenia y afines
[editar]|
Las gleicheniáceas son muy fáciles de distinguir debido a sus características frondes divididas pseudodicotómicamente ("falsamente divididas en dos"), lo que es logrado cuando las yemas apicales luego de dar las dos ramas laterales entran en dormición. Son terrestres de hojas coriáceas ("de aspecto de cuero"), a diferencia de muchos helechos no se encuentran en climas tropicales, solo en climas templados a templado-fríos. Carácter de importancia taxonómica: Poseen leptosporangios con anillo entero (no interrumpido en ninguna parte de su recorrido) oblicuo. |
 |
Schizaea, Lygodium, Anemia y afines
[editar]|
Esta familia es muy antigua y con un largo registro fósil. Sus 3 géneros más sobresalientes (Schizaea, Anemia y Lygodium) difieren mucho entre sí en la morfología del esporófito. Anemia es fácilmente reconocida porque las pinas fértiles son las basales y son muy reducidas, teniendo el aspecto de racimos dorados (por eso en Sudamérica subtropical se la conoce como "doradilla"). Schizaea tiene la particularidad de tener todas sus frondes reducidas ("filiformes"). Lygodium es conocido por ser un helecho trepador, muy apreciado en los jardines de clima cálido de todo el mundo. Lygodium japonicum es un helecho trepador cultivado como ornamental, también considerado invasor en Estados Unidos. Carácter de importancia taxonómica: El anillo de las schizáceas es entero, de posición transversal subapical, considerado un carácter primitivo dentro de los leptosporangios con anillo.
|
 |

Marsilea, Salvinia, Azolla y afines
[editar]|
Prácticamente todos los helechos acuáticos (salvo el género Ceratopteris que es un "helecho típico") pertenecen a este grupo. Marsilea es fácilmente confundida con el "trébol de 4 hojas", Salvinia es un conocido invasor de diques y aguas estancadas, y Azolla es cultivada en los campos de arroz por su poder fertilizante, adquirido en su relación simbiótica con una bacteria fijadora de nitrógeno. Su característica más sobresaliente es que poseen un ciclo de vida heterospórico. Carácter de importancia taxonómica: Poseen leptosporangios sin anillo o con anillo relictual (débilmente marcado, no funcional), por lo que fue perdido en forma secundaria (después de haberse adquirido).
|



Cyathea, Dicksonia y afines
[editar]|
Fácilmente reconocibles debido a su característico tallo formando un estípite que puede llegar a tener varios metros de altura, lo que le da al esporófito un aspecto arbóreo. Casi todos los helechos arborescentes pertenecen a este grupo. Normalmente se los encuentra divididos en varias familias, las más conspicuas son Cyatheaceae y Dicksoniaceae, la diferencia principal es que en las cyatheáceas hay haces vasculares en la médula del tallo (se dice que "la médula está revitalizada"). Carácter de importancia taxonómica: Poseen leptosporangios con anillo vertical completo (no interrumpido en ninguna parte de su recorrido). Según algunos autores (Tyron) debería llamarse anillo oblicuo.
|
 |

Helechos típicos
[editar]|
Comprende al 80 % de las especies de helechos. Normalmente se los encuentra divididos en una cantidad considerable de familias, entre las que se encuentran Pteridaceae, Aspleniaceae, Polypodiaceae, entre otras. Desde el 2004 (Pryer et al.), se sabe que son monofiléticos. Carácter de importancia taxonómica: Todas las especies tienen leptosporangio con anillo vertical incompleto, con estomio. Al deshidratarse el esporangio, el anillo se retrae y el estomio cede liberando las esporas de forma brusca, eso asegura una mejor dispersión de la especie con respecto a aquellas que tienen anillos sin estomio. Por ello se considera que este tipo de leptosporangio es el más avanzado. |
 |
 |
Taxonomía
[editar]- Introducción teórica en Taxonomía
La clasificación más actualizada es la de Christenhusz et al. 2011[1][2][3] (basada en Smith et al. 2006,[4] 2008);[5] que también provee una secuencia lineal de las licofitas y monilofitas.
Subclase: V Polypodiidae Cronquist, Takht. & Zimmerm., Taxon 15: 133 (1966). Sinónimos: Pterididae Schmakov, Turczaninowia 4: 42 (2001). Dennstaedtiidae Doweld, Tent. Syst. Pl. Vasc. (Tracheophyta): 10 (2001). Marsileidae Doweld, Tent. Syst. Pl. Vasc. (Tracheophyta): 10 (2001). Plagiogyriidae Doweld, Tent. Syst. Pl. Vasc. (Tracheophyta): 12 (2001).
Osmundales
[editar]Orden H. Osmundales Link, Hort. Berol. 2: 138 (1833). 1 familia:
- Familia 8. Osmundaceae Martinov, Tekhno-Bot. Slovar : 445 (1820).
- 4 géneros (Leptopteris, Osmunda, Osmundastrum, Todea). Referencias: Metzgar et al. (2008), Yatabe et al. (1999).
Hymenophyllales
[editar]Orden I. Hymenophyllales A.B. Frank en J.Leunis, Syn. Pflanzenk., ed. 2, 3: 1452 (1877). 1 familia.
- Familia 9. Hymenophyllaceae Mart., Consp. Regni Veg.: 3 (1835). Sinónimos: Trichomanaceae Burmeist., Handb. Naturgesch.: 196 (1836).
- 2 o más géneros. Referencias: Dubuisson (1996, 1997), Dubuisson et al. (2003), Ebihara et al. (2002, 2006, 2007), Hennequin et al. (2003, 2008).
- Nota: La familia consiste en 2 clados principales que se corresponden con los clásicos géneros Hymenophyllum y Trichomanes. El último es dividido en 8 géneros (Ebihara et al. 2006): Abrodictyum, Callistopteris, Cephalomanes, Crepidomanes, Didymoglossum, Polyphlebium, Trichomanes y Vandenboschia.
Gleicheniales
[editar]Orden J. Gleicheniales Schimp., Traité Paléont. Vég. 1: 669 (1869). Sinónimos: Matoniales Pic.Serm. ex Reveal, Phytologia 74: 175 (1993). Stromatoperidales Pic.Serm. ex Reveal, Phytologia 74: 176 (1993). Dipteridales Doweld, Tent. Syst. Pl. Vasc. (Tracheophyta): 10 (2001). 3 familias:
- Familia 10. Gleicheniaceae C.Presl, Reliq. Haenk.: 1: 70 (1825). Sinónimo: Stromatopteridaceae Bierh., Phytomorphology 18: 263 (1968).
- 6 géneros (Dicranopteris, Diplopterygium, Gleichenella, Gleichenia, Sticherus, Stromatopteris).
- Familia 11. Dipteridaceae Seward & E.Dale, Philos. Trans., ser. B 194: 487 (1901). Sinónimo: Cheiropleuriaceae Nakai, Bot. Mag. (Tokyo) 42: 210 (1928).
- 2 géneros (Cheiropleuria, Dipteris) Referencias: Kato et al. (2001).
- Familia 12. Matoniaceae C.Presl, Gefässbündel Farrn: 32 (1847).
- 2 géneros (Matonia, Phanerosorus). Referencias: Kato & Setoguchi (1998).
Schizaeales
[editar]Orden K. Schizaeales Schimp., Traité Paléonnt. Vég. 1: 674 (1869). 3 familias. Referencias: Dettmann & Clifford (1992), Skog et al. (2002), Wikström et al. (2002).
- Familia 13. Lygodiaceae M.Roem., Handb. Allg. Bot. 3: 520 (1840). 1 género (Lygodium). Referencia: Madeira et al. (2008).
- Familia 14. Schizaeaceae Kaulf., Wesen Farrenkr.: [119] (1827). 2 géneros (Actinostachys, Schizaea). Referencia: Wikström et al. (2002).
- Familia 15. Anemiaceae Link, Fil. Spec.: 23 (1841). 1 género (Anemia).
- Nota: Mohria está incluido en Anemia (Wikström et al. 2002).
Salviniales
[editar]Orden L. Salviniales Bartl. en Mart., Consp. Regn. Veg.: 4 (1835). Sinónimos: Pilulariales Bercht. & J.Presl, Přir. Rostlin : 272 (1820). Marsileales Bartl. en Mart., Consp. Regn. Veg.: 4 (1835). 2 familias:
- Familia 16. Marsileaceae Mirb. in Lam. & Mirb., Hist. Nat. Vég. 5: 126 (1802). Sinónimo: Pilulariaceae Mirb. ex DC., Essai Propr. Méd. Pl.: 48 (1804).
- 3 géneros (Marsilea, Pilularia, Regnellidium). Referencias: Nagalingum et al. (2008), Pryer (1999), Pryer & Hearn (2009), Schneider & Pryer (2002).
- Familia 17. Salviniaceae Martinov, Tekhno-Bot. Slovar: 559 (1820). Sinónimo: Azollaceae Wettst., Handb. Syst. Bot. 2: 77 (1903).
Cyatheales
[editar]Orden M. Cyatheales A.B.Frank en J.Leunis, Syn. Pflanzenk., ed. 2, 3: 1452 (1877). Sinónimos: Dicksoniales Pic.Serm. ex Reveal, Phytologia 74: 175 (1993). Hymenophyllopsidales Pic.Serm. ex Reveal, Phytologia 74: 175 (1993). Loxsomatales Pic.Serm. ex Reveal, Phytologia 74: 175 (1993). Plagiogyriales Pic.Serm. ex Reveal, Phytologia 74: 176 (1993). Metaxyales Doweld, Tent. Syst. Pl. Vasc. (Tracheophyta): 12. (2001). 8 familias. Referencias: Holttum & Sen (1961), Korall et al. (2006a).
- Familia 18. Thyrsopteridaceae C.Presl, Gefässbündel Farrn: 22, 38 (1847), como ‘Thyrsopterideae’.
- 1 género (Thyrsopteris). Referencias: Boodle (1915), Sen & Rahaman (1999).
- Familia 19. Loxsomataceae C.Presl, Gefässbündel Farrn: 31 (1847), como ‘Loxsomaceae’.
- 2 géneros. (Loxsoma , Loxsomopsis). Referencias: Bower (1923), Lehnert et al. (2001).
- Nota: El nombre ‘Loxoma’ fue un error de ortografía y fue corregido a Loxsoma (Hooker 1838) .
- Familia 20. Culcitaceae Pic.Serm., Webbia 24: 702 (1970).
- 1 género (Culcita). Referencia: Sen (1968).
- Familia 21. Plagiogyriaceae Bower, Ann. Bot. (London) 40: 484 (1926).
- 1 género (Plagiogyria). Referencia: Zhang & Nooteboom (1998).
- Familia 22. Cibotiaceae Korall in A.R.Sm. et al., Taxon 55: 712 (2006).
- 1 género (Cibotium). Referencia: Smith et al. (2006a).
- Familia 23. Cyatheaceae Kaulf., Wesen Farrenkr.: [119] (1827). Sinónimos: Alsophilaceae C.Presl, Gefässbündel Farrn: 22 (1847). Hymenophyllopsidaceae Pic.Serm., Webbia 24: 712 (1970).
- 4 géneros ( Alsophila, Cyathea, Gymnosphaera, Sphaeropteris). Referencias: Christenhusz (2009b), Conant et al. (1995, 1996), Conant & Stein (2001), Janssen et al. (2008), Korall et al. (2006a), Lantz et al. (1999), Lehnert (2006, 2008, 2009), Marquez (2010), Stein et al. (1996), Wang et al. (2003) .
- Familia 24. Dicksoniaceae M.R.Schomb., Reis. Br.-Guiana (Ri. Schomburgk) 2: 1047 (1848). Sinónimo: Lophosoriaceae Pic.Serm., Webbia 24: 700 (1970).
- 3 géneros (Calochlaena, Dicksonia, Lophosoria). Referencias: Churchill et al. (1998), Lehnert (2006), White & Turner (1988).
- Familia 25. Metaxyaceae Pic.Serm., Webbia 24: 701 (1970).
- 1 género (Metaxya). Referencias: Qiu et al. (1995), Sen (1969), Smith et al. (2001).
Polypodiales
[editar]Orden N. Polypodiales Link, Hort. Berol. 2: 5 (1833). Sinónimos: Parkeriales A.B.Frank en J.Leunis, Syn. Pflanzenk., ed. 2, 3: 1452 (1877). Filicales Dumortier, Anal. Fam. Pl.: 67. 1829, nom. illeg. Blechnales Pic.Serm. ex Reveal, Phytologia 74: 175 (1993). Negripteridales Pic.Serm. ex Reveal, Phytologia 74: 176 (1993). Platyzomatales Pic.Serm. ex Reveal, Phytologia 74: 176 (1993). Aspleniales Pic.Serm. ex Reveal, Phytologia 79: 72 (1996) Athyriales Schmakov, Turczaninowia 4: 55 (2001). Dennstaedtiales Doweld, Tent. Syst. Pl. Vasc. (Tracheophyta): 10 (2001). Pteridales Doweld, Tent. Syst. Pl. Vasc. (Tracheophyta): 11 (2001). Dryopteridales Schmakov en Turczaninowia 4: 66 (2001). Thelypteridales Doweld, Tent. Syst. Pl. Vasc. (Tracheophyta): 11 (2001). Lindsaeales Doweld, New Syllabus: 353 (2006).
21 familias:
- Familia 26. Lonchitidaceae C.Presl ex M.R.Schomb., Reis. Br.-Guiana (Ri. Schomburgk) 2: 1047 (1848). 1 género (Lonchitis). Referencias: Christenhusz (2009a), Lehtonen et al. (2010), Lellinger (1977).
- Familia 27. Saccolomataceae Doweld en Doweld & Reveal, Phytologia 90: 417 (2008). 1–2 géneros (Orthiopteris, Saccoloma). Referencia: Nair (1992).
- Nota: El estado de Orthiopteris como separado de Saccoloma todavía es incierto. Se necesitan más estudios filogenéticos de la familia.
- Familia 28. Cystodiaceae J.R.Croft, Kew Bull. 41: 797 (1986). 1 género (Cystodium). Referencias: Croft (1986), Korall et al. (2006b), Lehtonen et al. (2010).
- Familia 29. Lindsaeaceae C.Presl ex M.R.Schomb., Reis. Br.-Guiana (Ri. Schomburgk) 2: 883 (1848). 7 géneros (Lindsaea, Nesolindsaea, Odontosoria, Osmolindsaea, Sphenomeris, Tapeinidium, Xyropteris). Referencias: Kramer (1957, 1967a, b, c, 1970, 1971, 1972a, b, 1989), Kramer & Tindale (1976), Lehtonen et al. (2010), Lin et al. (1999), Schneider & Kenrick (2001).
- Familia 30. Dennstaedtiaceae Lotsy, Vortr. Bot. Stammesgesch. 2: 655 (1909). Sinónimos: Hypolepidaceae Pic.Serm., Webbia 24: 705 (1970). Pteridiaceae Ching, Acta Phytotax. Sin. 13: 96 (1975). 10 géneros (Blotiella, Dennstaedtia, Histiopteris, Hypolepis, Leptolepia, Microlepia, Monachosorum, Oenotrichia, Paesia, Pteridium). Referencias: Der et al. (2009), Wolf (1995). Nota: Oenotrichia está basado en O. maxima (E.Fourn.) Copel., que pertenece a Dennstaedtiaceae. Las otras especies pertenecen a Dryopteridaceae.
- Familia 31. Pteridaceae E.D.M.Kirchn., Schul-Bot.: 109 (1831). Sinónimos: Parkeriaceae Hook., Exot. Fl. 2: adt. 147 (1825), nom. rej. en favor de Adiantaceae Newman (1840). Adiantaceae Newman, Hist. Brit. Ferns: 5 (1840), nom. cons. Acrostichaceae Mett. ex A.B.Frank en J.Leunis, Syn. Pflanzenk., ed. 2, 3: 1453 (1874). Ceratopteridaceae Underw., Our Native Ferns, ed. 6: 65 (1900). Sinopteridaceae Koidz., Acta Phytotax. Geobot. 3: 50 (1934). Vittariaceae Ching, Sunyatsenia 5: 210 (1940). Negripteridaceae Pic.Serm., Nuovo Giorn. Bot. Ital., ser. 2, 53: 160 (1946). Platyzomataceae Nakai, Bull. Natl. Sci. Mus. Tokyo 29: 4 (1950). Actiniopteridaceae Pic.Serm., Webbia 17: 5 (1962). Cryptogrammaceae Pic.Serm., Webbia 17: 299 (1963). Hemionitidaceae Pic.Serm., Webbia 21: 487 (1966). Cheilanthaceae B.K.Nayar, Taxon 19: 233 (1970). Taenitidaceae Pic.Serm., Webbia 29: 1 (1975). Antrophyaceae Ching, Acta Phytotax. Sin. 16: 11 (1978). Anopteridaceae Doweld, Tent. Syst. Pl. Vasc. (Tracheophyta): 2 (2001). Géneros: 50–60 (53 aceptados aquí). Referencias: Beck et al. (2010), Gastony & Johnson (2001), Gastony & Rollo (1995, 1998), Grusz et al. (2009), Kirkpatrick (2007), Nakazato & Gastony (2001), Prado et al. (2007), Rohfelds et al. (2008), Ruhfel et al. (2008), Sánchez-Barracaldo (2004), Schuettpelz et al. (2007), Windham et al. (2009), Zhang et al. (2005, 2007).
- Subfamilia 31a. Cryptogrammoideae S.Linds. Edinburgh J. Bot. 66(2): 358. 2009 (Coniogramme, Cryptogramma, Llavea).
- Subfamilia 31b. Ceratopteridoideae (J.Sm.) R.M.Tryon, Amer. Fern J. 76: 184 (1986). Sinónimo: ‘Parkerioideae’ de Smith et al. (2006a), nom. nud. (Acrostichum, Ceratopteris ).
- Subfamilia 31c. Pteridoideae C.Chr. ex Crabbe, Jermy & Mickel, Fern Gaz. 11: 153 (1975). Sinónimo: Taenitidoideae (C.Presl) R.M.Tryon, Amer. Fern J. 76: 184 (1986). (Actiniopteris, Anogramma, Aspleniopsis, Austrogramme, Cerosora, Cosentinia, Jamesonia, Nephopteris, Onychium, Pityrogramma, Platyzoma, Pteris, Pterozonium, Syngramma, Taenitis) Nota: Eriosorus está unido con Jamesonia (Sánchez-Barracaldo 2004) y Neurocallis y Platyzoma están unidos a Pteris (Schuetpeltz & Pryer 2007).
- Subfamilia 31d. Cheilanthoideae W.C.Shieh, J. Sci. Engin. (Nation. Chung-Hsing Univ.) 10: 211 (1973). (Adiantopsis, Aleuritopteris, Argyrochosma, Aspidotis, Astrolepis, Bommeria, Calciphilopteris, Cassebeera, Cheilanthes, Cheiloplecton, Doryopteris, Hemionitis, Mildella, Notholaena, Paraceterach, Paragymnopteris, Pellaea, Pentagramma, Trachypteris, Tryonella). Nota: La clasificación de géneros de helechos de la familia Cheilanthoideae no es estable porque muchos de los conceptos de género no comprenden unidades naturales, por ejemplo, Paraceterach como clasificada por Tryon et al. (1990) es parafilético (Kirkpatrick 2007) y embebido en Pellaea s.l. Los helechos cheilantoides están bajo estudio ahora en el laboratorio de "Pryer lab" (Beck et al. 2010, Grusz et al. 2009, Rothfels et al. 2008, Windham et al. 2009).
- Subfamilia 31e. Vittarioideae (C.Presl) Crabbe, Jermy & Mickel, Fern Gaz. 11: 154 (1975). Sinónimo: Adiantoideae (C.Presl) R.M.Tryon, Amer. Fern J. 76: 184 (1986). (Adiantum, Ananthacorus, Anetium, Antrophyum, Haplopteris, Hecistopteris, Monogramma, Polytaenium, Radiovittaria, Rheopteris, Scoliosorus, Vittaria). Nota: Los helechos vittarioides están embebidos en el género Adiantum, por eso aquí son tratados en una única subfamilia. Adiantum necesita revisión taxonómica.
- Familia 32. Cystopteridaceae Schmakov, Turczaninowia 4: 60 (2001). 4 géneros. (Acystopteris, Cystoathyrium, Cystopteris, Gymnocarpium).
- Descripción: Plantas de tamaño pequeño o mediano, en bosques o en grietas. Rizoma esbelto largamente rastrero, o cortamente rastrero, o cortos erectos o ascendentes; lámina 1-3 pinada; venas libres; soros pequeños, redondeados a forma de copa, dorsales en las venillas; indusios pequeños, delgados aovados o ovales, adjuntados en las bases, como escamas e inmersos en los esporangios maduros, o sin indusio; x=42, 41, 40.
- Nota:—Schuettpelz & Pryer (2007) encontraron que Gymnocarpium y Cystopteris son hermanos del resto de eupolipoides II. Por lo tanto esos géneros y sus afines Acystopteris y Cystoathyrium son ubicados aquí en su propia familia, en sentido filogenético.
- Familia 32.1? Hemidictyaceae, fam. nov. Descripción: Frondes herbaceae, compositae, pinnis oppositis lanceolatis serratis; fasciculus vasorum in stipite unicus; venae pinnatae, simplices; sori lineares, elongati, dorso venae simplicis; indusium lineare, elongatum, planum. Tipo: Hemidictyum C.Presl (1836: 110)] 1 género (Hemidictyum)
- Nota: Este género podría ser incluido en Aspleniaceae (como sugerido por Lehtonen 2011), si bien es indeseable porque las sinapomorfías de un Aspleniaceae definido de forma tan amplia aún no son conocidas. Más aún, la separación de Hemidictyum y Aspleniaceae data del Cretácico de acuerdo con las estimaciones actuales de divergencia de los helechos derivados (Schuettpelz & Pryer 2009), y el reconocimiento de dos familias sería por lo tanto buena ayuda para la clasificación de helechos del Cretácico y del Terciario temprano.
- Familia 33. Aspleniaceae Newman, Hist. Brit. Ferns: 6 (1840). 2 géneros. (Asplenium, Hymenasplenium). Referencias: Murakami & Schaal (1994), Murakami et al. (1999), Perrie & Brownsey (2005), Pinter al. (2002), Schneider et al. (2004b, 2005).
- Nota: Los estudios citados arriba han encontrado a todos los segregados de Asplenium (por ejemplo Camptosorus, Ceterach, Diellia, Loxoscaphe y Phyllitis) anidados dentro de él. Además se conocen muchos híbridos entre géneros. Por eso Smith et al. (2006a) reconocieron solo dos géneros en Aspleniaceae que se segregan por citología, y es el tratamiento que se sigue aquí.
- Familia 34. Diplaziopsidaceae X.C.Zhang & Christenh., fam. nov. 2 géneros (Diplaziopsis (tipo de la familia), Homalosorus). En la primera versión del escrito se encontraba Hemidictyum que en la corrección se ubica en su propia familia Hemidictyaceae. Filices mediocres vel grandes; rhizomatibus crassis, erectis vel decumbentibus; laminis 1-pinnatis, imparipinnatis, pinnis integris, glabris; venis lateralibus utroque costulae latere discretis sed margines versus reticulatim anastomosantibus, areolarum 2–4 series facientibus, sine ven ulis discretis inclusis; soris secus venas longas prope costulam elongatis, indusio similiter elongato membranaceo obtectis; indusiis plerumque 3–8 mm, interdum ad 1–2 cm longis, arcuatis, allantoideis; número básico chromosomatum x=31, 40, 41. Genus quod est typus familiae: Diplaziopsis C.Chr.
- Descripción: Helechos de tamaño mediano a grande, usualmente en bosques cerca de o en corrientes de agua. Rizomas decumbentes a erectos, gruesos, lámina de 1 pina, imparipinada, con pinas enteras glabras; venas libres en los dos lados de la vena media, pero anastomosándose totalmente hacia los márgenes donde forman 2 a 4 filas de areolas, sin incluir las venillas libres; soros elongándose a lo largo de las venas grandes cerca de la vena madia, cubiertos por indusio similarmente elongado y membranoso; indusios usualmente de 3 a 8 mm, a veces hasta 1-2 cm de largo, arqueados, como salchichas; esporas con pliegues como alas fimbriadas y "echinate" (en inglés), o con pliegues perforados en la parte baja. X=41, 40 or 31.
- Notas: Homalosorus pycnocarpos (Spreng.) Pic.Serm. ha sido transferido a Diplaziopsis por Price (1990). Diferente del asiático Diplaziopsis con x=41, el norteamericano templado Homalosorus pycnocarpos tiene x=40 y el género tropical americano Hemidictyum marginatum tiene x=31. Hemidictyum primero fue mostrado que era hermano de Aspleniaceae por Schuetpelz & Pryer (2007), pero Diplaziopsis no fue incluido en su estudio. Aquí incorporamos evidencia del laboratorio de X.-C. Zhang, y ubicamos a estos géneros en una familia (Wei et al. 2010). La inclusión de Hemidictyum en Diplaziopsidaceae es sin embargo todavía tentativa; más estudios se necesitan para confirmar esta ubicación.
- Familia 35. Thelypteridaceae Pic.Serm., Webbia 24: 709 (1970). 5 o más géneros. Referencias: Smith (1986), Smith & Cranfill (2002), Yatabe et al. (2002).
- Nota: Cinco géneros (Cyclosorus, Macrothelypteris, Phegopteris, Pseudophegopteris, Thelypteris) fueron reconocidos por Smith (en Kramer & Green 1990) y muchos géneros antiguos fueron incluidos en Cyclosorus y Thelypteris. La comprensión que se tiene en estos momentos de la filogenia de la familia es limitada por lo que utilizamos una aproximación conservativa, con conceptos de género amplios, para las especies que pertenecen a Cyclosorus y Thelypteris. Conceptos de género más estrechos son discutidos para reemplazarlos por estos dos grandes géneros pero la evidencia actual es todavía insuficiente para llegar a una conclusión final.
- Familia 36. Woodsiaceae Herter, Rev. Sudamer. Bot. 9: 14 (1949). 1–3 géneros (Cheilanthopsis, Hymenocystis, Woodsia). Notas: Woodsiaceae antes incluía a Athyriaceae y Diplaziopsidaceae. La familia aquí es definida en su sentido más estricto, incluyendo solo a Woodsia y dos géneros altamente relacionados que, a la espera de más evidencia, quizás terminen siendo unidos a Woodsia en el futuro. Los linajes de Athyriaceae, Cystopteridaceae, Diplaziopsidaceae y Rachidosoraceae están consistentemente segregados en todos los análisis, independientemente del muestreo. Si bien las relaciones entre estos grupos no siempre quedan resueltas, estos linajes no pueden ser unidos a otros más grandes. Por eso son tratados en familias separadas.
- Familia 37. Rhachidosoraceae X.C.Zhang, fam. nov. Basada en una referencia completa y directa de la descripción latina asociada con Athyriaceae subfam. Rhachidosoroideae M.L.Wang & Y.T.Hsieh, Acta Phytotax. Sin. 42: 527 (2004). 1 género. (Rhachidosorus Ching, tipo de la familia).
- Descripción: Helechos medianos a grandes en bosques muchas veces en lugares rocosos (piedra caliza). Rizomas gruesos, erectos a decumbentes, los ápices y bases de los tallos densamente cubiertos por escamas, láminas 2-3pinadas, deltoides a aovadas-deltoides, soros lineales, falcados, que tocan a las venas medias en el extremo proximal, subparalelos a la vena media. Indusios más o menos gruesos, enteros, en el lado acroscópico de las venillas laterales, asplenioides, esporas con pliegues tuberculares. X=40. Un solo género con cerca de 7 especies, distribuido en el este y el sudeste de Asia, desde Japón a Sumatra y las Filipinas. Rhachidosorus no fue incluido en el estudio de Schuettpelz & Pryer (2007) y los resultados de Wang et al. (2003, 2004) no fueron aceptados por Smith et al. (2006a, 2008).
- Familia 38. Onocleaceae Pic.Serm., Webbia 24: 708 (1970). 1(–4) géneros: (Onoclea). Referencia: Gastony & Ungerer (1997).
- Nota: Los cuatro géneros (Matteuccia, Onoclea, Onocleopsis y Pentarhizidium) son muy cercanos por lo que aquí son tratados como un único género, Onoclea.
- Familia 39. Blechnaceae Newman, Hist. Brit. Ferns, ed. 2: 8 (1844). Sinónimo: Stenochlaenaceae Ching, Acta Phytotax. Sin. 16: 18 (1978). 2–9 géneros. (Blechnum, Brainea, Pteridoblechnum, Sadleria, Salpichlaena, Stenochlaena, Woodwardia). Referencias: Nakahira (2000), Cranfill (2001), Cranfill & Kato (2003). Nota: El género Doodia está incluido en Blechnum (Shepherd et al. 2007). Blechnum en su circunscripción actual, aun incluyendo a Doodia, probablemente sea parafilético. El estado de otros géneros como Brainea, Pteridoblechnum, Sadleria, Salpichlaena y Stenochlaena aún no está claro, pero algunos de éstos probablemente deban ser incluidos en Blechnum sensu lato (ver Cranfill 2001). Se necesitan más estudios en la delimitación de los géneros de esta familia.
- Familia 40. Athyriaceae Alston, Taxon 5: 25 (1956). 5 géneros. (Anisocampium, Athyrium, Cornopteris, Deparia, Diplazium) Referencias: Kato (1977), Wang et al. (2004).
- Nota: Athyriaceae incluye a la mayoría de los géneros ubicados por Smith et al. (2006a, 2008) en Woodsiaceae. La clasificación de los géneros necesita más estudios, especialmente en el contexto de la monofilia de Athyrium y Diplazium.
- Familia 41. Hypodematiaceae Ching, Acta Phytotax. Sin. 13: 96 (1975). 3 géneros (Didymochlaena, Hypodematium, Leucostegia). Referencias: Liu et al. (2007a), Tsutsumi & Kato (2006), Schuettpelz & Pryer (2007).
- Nota: Estos géneros tradicionalmente no fueron asociados. Hypodematium fue asociado con los helechos athirioides, Didymochlaena fue antes ubicado en Dryopteridaceae y Leucostegia en Davalliaceae. Muchos estudios (Liu et al. 2007a, Tsutsumi & Kato 2006, Schuettpelz & Pryer 2007) han encontrado que están relacionados y probablemente son el clado hermano de 'Eupolypods I'.
- Familia 42. Dryopteridaceae Herter, Rev. Sudamer. Bot. 9: 15 (1949), nom. cons. Sinónimos: Aspidiaceae Mett. ex A.B.Frank en Leunis, Syn. Pflanzenk. ed. 2. 3: 1469 (1877), nom. illeg. Filicaceae Juss., Gen. Pl.: 14. (1789), como 'Filices', nom. illeg., rej. Peranemataceae Ching, Sunyatsenia 5: 208 (1940), nom. rej. Elaphoglossaceae Pic.Serm., Webbia 23: 209 (1968). Bolbitidaceae Ching, Acta Phytotax. Sin. 16: 15 (1978). Cerca de 34 géneros. Referencias: Li & Lu (2006), Li et al. (2008), Liu et al. (2007a, 2010), Moran et al. (2010a, b), Rouhan (2004). Nota: Son necesarios más estudios a nivel de género para la ubicación correcta de los géneros y para confirmar la monofilia de los géneros ricos en especies Dryopteris y Polystichum. Insertis sedis: Adenoderris, Coveniella, Dracoglossum, Revwattsia, Stenolepia.
- Subfamilia 42a. Dryopteridoideae B.K.Nayar, Taxon 19: 235 (1970). (Acrophorus, Acrorumohra, Arachniodes, Ctenitis, Cyrtogonellum, Cyrtomidictyum, Cyrtomium, Diacalpe, Dryopolystichum, Dryopsis, Dryopteris, Leptorumohra, Lithostegia, Peranema, Phanerophlebia, Polystichopsis, Polystichum).
- Subfamilia 42b. Elaphoglossoideae (Pic.Serm.) Crabbe, Jermy & Mickel, Fern Gaz. 11: 154 (1975). Basado en Elaphoglossaceae Pic.Serm. (Arthrobotrya, Bolbitis, Cyclodium, Elaphoglossum, Lastreopsis, Lomagramma, Maxonia, Megalastrum, Mickelia, Olfersia, Polybotrya, Rumohra, Stigmatopteris, Teratophyllum).
- Familia 43. Lomariopsidaceae Alston, Taxon 5: 25 (1956). 3 géneros (Cyclopeltis, Lomariopsis, Thysanosoria). Referencias: Moran (2000), Rouhan et al. (2007), Tsutsumi & Kato (2006), Schuettpelz & Pryer (2007), Liu et al. (2007a).
- Familia 44. Nephrolepidaceae Pic.Serm., Webbia 29: 8 (1975). 1 género (Nephrolepis). Referencias: Hennequin et al. (2010), Hovenkamp & Miyamoto (2005). Nota: El género Nephrolepis siempre fue dificultoso de ubicar. Ha sido antes asociado a Davalliaceae y a Oleandraceae (Kramer & Green 1990). Smith (2006a, 2008) lo ubica en Lomariopsidaceae, que comparte con él las pinas articuladas. Esta asociación no es satisfactoria sin embargo porque su exacta ubicación filogenética todavía es incierta. Por eso nosotros lo ubicamos tentativamente en su propia familia hasta que haya más datos disponibles.
- Familia 45. Tectariaceae Panigrahi, J. Orissa Bot. Soc. 8: 41 (1986). Sinónimos: Dictyoxiphiaceae Ching, Sunyatsenia 5: 205, 218. 1940, nom. inval. Hypoderriaceae Ching, Sunyatsenia 5: 209, 245. 1940, nom. inval. 6–10 géneros. (Aenigmopteris, Arthropteris, Hypoderris, Pleocnemia, Psammiosorus, Psomiocarpa, Pteridrys, Tectaria, Triplophyllum, Wagneriopteris). Referencias: Christenhusz (2010b), Liu (2007b).
- Nota: Arthropteris ha sido tentativamente incluido aquí por Smith et al. (2006a), pero Liu (2007b) lo ubicó en Dryopteridaceae. La correcta ubicación de este género todavía es incierta. Esta familia está pobremente definida, especialmente cuando son incluidos Arthropteris y Psammiosorus. El número de géneros que pertenecen a Tectariaceae todavía es incierto.
- Familia 46. Oleandraceae Ching ex Pic.Serm., Webbia 20: 745 (1965). 1 género (Oleandra). Referencias: Tryon (1998, 2000)
- Familia 47. Davalliaceae M.R.Schomb., Reis. Br.-Guiana (Ri. Schomburgk) 2: 883 (1848). 2 géneros (Davallia, Davallodes). Referencia: Kato & Tsutsumi (2009), Tsutsumi & Kato (2006), Tsutsumi et al. (2008).
- Nota: Tsutsumi et al. (2006, 2008) proveyeron una filogenia de Davalliaceae, donde encontraron que Davallia es polifilético y propusieron reconocer los cinco clados a nivel de género. Esta clasificación aumenta mucho el número de géneros y un concepto alternativo con solo dos géneros también puede ser considerado. Propusieron al género Araiostegiella Kato & Tsutsumi (2008) y reinstauraron Wibelia Bernhardi (1801: 122), el último terminó siendo un homónimo de Wibelia Gaertner et al. (1801: 97, 144), un género de Asteraceae (= Crepis), por lo que un nuevo nombre de género es necesario para ubicar a Wibelia Bernh. La mayoría de las especies tiene combinaciones con Davallia y parece apropiado considerar un concepto más amplio del género Davallia que incluya a Araiostegiella y a Humata. Para dividir a Davallia, la pregunta es a cuál de los 5 clados el nombre Davallia debería ser aplicado, porque la geográficamente aislada D. canariensis (L.) Sm., la especie tipo, no está incluida en estos estudios. Sin establecer a qué clado pertenece esta especie, una subdivisión de géneros es difícil de validar. Davallodes (incl. Araiostegia) como tratado por Tsutsumi et al. (2008) es utilizado aquí, reduciendo el número de géneros a dos, pero esto puede necesitar ser actualizado con un mejorado análisis filogenético en el futuro.
- Familia 48. Polypodiaceae J.Presl & C.Presl, Delic. Prag.: 159 (1822). Sinónimos: Grammitidaceae Newm., Hist. Brit. Ferns: 7 (1840). Gymnogrammitidaceae Ching en Acta Phytotax. Sin. 11: 12 (1966). Loxogrammaceae Ching ex Pic.Serm. en Webbia 29: 11 (1975). Drynariaceae Ching en Acta Phytotax. Sin. 16: 19 (1978). Platyceriaceae Ching en Acta Phytotax. Sin. 16: 18 (1978). Pleurisoriopsidaceae Kurita & Ikebe ex Ching en Acta Phytotax. Sin. 16: (1978). 50–80 géneros (64 aceptados aquí tentativamente). Referencias: Janssen & Schneider (2005), Kreier et al. (2008), Labiak et al. (2010a, b), Otto et al. (2009), Parris (2007), Ranker (2004), Schneider et al. (2004a, c, 2006a, b, 2008, 2010), Smith et al. (2006b), Wang et al. (2010a, b).
- Subfamilia 48a. Loxogrammoideae H.Schneid., subfam. nov. (Dictymia, Loxogramme). Basónimo: Loxogrammaceae Ching ex Pic. Serm., Webbia 29: 11 (1974). Tipo: Loxogramme.
- Subfamilia 48b. Drynarioideae Crabbe, Jermy & Mickel, Fern Gaz. 11: 156 (1975). (Aglaomorpha, Arthromeris, Christiopteris, Drynaria, Gymnogrammitis, Paraselliguea, Phymatopteris, Polypodiopteris, Selliguea). Nota: Esta familia incluye a los helechos drynarioides y selligueoides. El número de géneros de ambos grupos todavía es incierto. Solo reconocemos a los géneros que son ampliamente aceptados.
- Subfamilia 48c. Platycerioideae B.K.Nayar, Taxon 19: 233 (1970). (Platycerium, Pyrrosia ).
- Subfamilia 48d. Microsoroideae B.K.Nayar, Taxon 19: 233 (1970), como ‘Microsorioideae’. Sinónimo: Lepisoroideae Ching, Acta Phytotax. Sin. 16: 17 (1978), como ‘Lepisorioideae’. (Dendroconche, Goniophlebium, Kaulinia, Kontumia, Lecanopteris, Lemmaphyllum, Lepisorus, Lepidomicrosorium, Leptochilus, Microsorum, Neocheiropteris, Neolepisorus, Paragramma, Phymatosorus, Podosorus, Thylacopteris, Tricholepidium). Nota: El estado de muchos géneros de este grupo es controvertido (Kreier et al. 2008). El género Microsorum es polifilético y deberá ser dividido en géneros naturales en el futuro. El género monotípico Kontumia es tentativamente asignado a los helechos microsoroides y no a los selligueoides como sugerido por Wu et al. (2005). Esta decisión está basada en las escamas del rizoma observadas por P. Hovenkamp (Leiden) que estudió el material tipo. Actualmente nos faltan secuencias de ADN para confirmar esta relación.
- Subfamilia 48e. Polypodioideae B.K.Nayar, Taxon 19: 234 (1970). Sinónimo: Pleopeltidoideae B.K.Nayar, Taxon 19: 234 (1970). (Campyloneurum, Microgramma, Niphidium, Pecluma, Phlebodium, Pleopeltis, Pleurosoriopsis, Polypodium, Serpocaulon, Synammia, y los grammítidos: Acrosorus, Adenophorus, Calymmodon, Ceradenia, Chrysogrammitis, Cochlidium, Ctenopterella, Dasygrammitis, Enterosora, Grammitis, Lellingeria, Leucostrichum, Luisma, Melpomene, Micropolypodium, Oreogrammitis, Prosaptia (incl. Ctenopteris), Radiogrammitis, Scleroglossum (incl. Nematopteris), Terpsichore, Themelium, Tomophyllum, Xiphopterella, Xiphoteris, Zygophlebia). Nota: El número de géneros de los helechos grammítidos todavía no está fijo. Un grupo de investigación bajo el liderazgo del Dr. Tom Ranker actualmente está estudiando este complejo.
Clasificación sensu Smith et al. 2006
[editar]Clados y taxones superiores: Plantae (clado), Viridiplantae, Streptophyta, Streptophytina, Embryophyta, Tracheophyta, Euphyllophyta, Monilophyta, Clase Polypodiopsida.
Según Smith et al. 2006, es sinónimo de la antigua Filicopsida, ya que comprende a la mayor parte de las plantas que comúnmente se llaman "helechos". El sistema de clasificación de Engler coincidió bastante con esta delimitación de Filicopsida, pero dejó en ella también a las Marattiopsidas y a las Ophioglossaceae, que también son helechos con megafilos. Los sistemas de clasificación más modernos como el de Bold et al. 1989 ya coincidían con que los helechos leptosporangiados eran un clado y también los llamaron Filicopsida o Pteropsida (o Pterophyta si orden), dejando en otra clase a las ofioglosáceas y las maratiáceas, que son eusporangiadas.
Circunscripción: Basados en los resultados de filogenia, han sido consensuados 7 órdenes y 33 familias:
Árbol filogenético actualizado y taxonomía de las clases y órdenes de monilophytas. Dibujado a partir de Smith et al. (2006). |
"Núcleo de los leptosporangiados": Salviniales + Cyatheales + Polypodiales Órdenes y familias basados en los resultados de la filogenia:
 Árbol filogenético actualizado y taxonomía de Salviniales y Cyatheales en el "Núcleo de los leptosporangiados". Dibujado a partir de Smith et al. (2006). |
- Salviniales ("helechos acuáticos", "helechos heterospóricos")
- Polypodiales ("polypods"). Basados en los resultados de la filogenia:
Árbol filogenético actualizado y taxonomía de Polypodiales. Dibujado a partir de Smith et al. (2006). |
"Eupolypodiales"
"Eupolypodiales II" (Aspleniaceae + Woodsiaceae + Thelypteridaceae + Blechnaceae + Onocleaceae)
"Eupolypodiales I" (Dryopteridaceae + Lomariopsidaceae + Tectariaceae + Oleandraceae + Davalliaceae + Polypodiaceae)
Los órdenes están bien consensuados y muy probablemente ésta ya sea la clasificación definitiva de los mismos, muchas familias ya están consensuadas también, pero otras necesitan más análisis. En el caso particular de Pteridaceae, como definido por Smith et al. es monofilético, pero compuesto por 5 lineajes bien definidos, así que quizás en el futuro se lo divida en 5 familias o subfamilias. En el caso de los géneros, prácticamente todos necesitan revisión, en especial los de las familias de polypodiales, donde hay muchos géneros polifiléticos y muchos parafiléticos. Los análisis pertinentes ya se están haciendo.
Clasificación de Bold & al. (1989)
[editar]Filicopsida sensu Bold & et al. es equivalente a Polypodiidae y diferente del taxón mayor Filicopsida, el cual agrupa también a Marattiales, Psilotales y Ophioglossales.
- división Pteridophyta
- clase Filicopsida sensu Bold, dividida en los órdenes:
Según este sistema, que deja afuera a los helechos eusporangiados, e taxón comprende unos 300 géneros y 9000 especies, más abundantes en los trópicos, desde plantas pequeñas hasta helechos arborescentes, las familias se distinguen por la morfología de los leptoesporangios.
- Primofílices.
- Orden Osmundales
- Osmundaceas (familia Osmundaceae), osmundas
- Orden Polypodiales.
- Polipodiaceas (familia Polypodiaceae).
- Orden Pteridales.
- Sinopteridaceas (familia Sinopteridaceae).
- Criptogramaceas (familia Cryptogrammaceae).
- Pteridaceas (familia Pteridaceae).
- Adiantaceas (familia Adiantaceae).
- Hemionitidaceas (familia Hemionitidaceae).
- Orden Marsileales.
- Marsileaceas (familia Marsileaceae).
- Orden rigodium implexum '
- Himenofilaceas (familia Hymenophyllaceae).
- Orden Dicksoniales.
- Culcitaceas (familia Culcitaceae).
- Dicksoniaceas (familia Dicksoniaceae).
- Orden Dennstaedtiales.
- Hipolepidaceas (familia Hypolepidaceae
- Dennstaedtiaceas (familia Dennstaedtiaceae).
- Orden Aspidiales.
- Telipteridaceas (familia Thelypteridaceae).
- Aspleniaceas (familia Aspleniaceae).
- Woodsidaceas (familia Woodsidaceae).
- Atiriaceas (familia Athyriaceae).
- Aspidiaceas (familia Aspidiaceae).
- Davaliaceas (familia Davalliaceae).
- Orden Blechnales.
- Bleknaceas (familia Blechnaceae).
- Orden Salviniales, acuáticos.
- Azolaceas (familia Azollaceae).
- Salviniaceas (familia Salviniaceae).
Ecología
[editar]Terrestres, epífitas o acuáticas. Gran diversidad de hábitos y adaptaciones.
Evolución
[editar]Las polypodiopsidas posee un largo registro fósil, salvo su clado más conspicuo, las polypodiales, que comprenden cerca del 80 % de los helechos, y cuyo registro fósil se enriquece en el Terciario. Schneider et al. (2004) hipotetizan que este clado se diversificó de forma oportunista, aprovechando nuevos nichos ecológicos provistos por el nuevo ambiente dominado por angiospermas (Schneider et al. 2004).
Aumento de la diversidad de helechos polypodiales en relación al aumento de la diversidad de angiospermas, ocurrido durante el Cretácico. Se muestran dos estimaciones de la diversificación de las angiospermas, correspondientes a la estimación según cantidad de bases de ADN cambiadas, corregida con respecto a la edad de los fósiles encontrados pertenecientes a angiospermas (la estimación de la derecha muestra una aplicación "estricta" de la edad de los fósiles de angiospermas, fijada en 132 millones de años, la de la izquierda muestra una aplicación "relajada" de la edad de los fósiles de angiospermas, en la que 132 millones de años es el valor mínimo posible). Dibujado y traducido a partir de Schneider et al. 2004. |
Caracteres
[editar]Xilema primario con placas de perforación escalariforme.
Los tallos son desde rizomatosos a arborescentes, que pueden llegar hasta los 20 metros de altura.
Las hojas son megafilos, usualmente grandes (bastante más de 2 cm de largo), y con numerosas venas bifurcadas ("forked"). En general las láminas de las hojas están divididas en lóbulos o pinas, a veces varias veces divididas, pero también se pueden encontrar hojas de lámina entera. Las hojas están dispuetas en espiral alrededor del tallo, a veces "2-ranked". Los pecíolos de las hojas suelen ser persistentes, por lo que quedan en el tallo una vez muerta la hoja, a veces cubriéndolo de una forma conspicua, pero también hay especies donde las hojas tienen una clara zona de abscisión por la que se desprenden del tallo en la senescencia (carácter habitual en especies epífitas). Las hojas pueden ser monomórficas (un solo morfo de hojas) o dimórficas (dos morfos de hojas diferentes en la misma planta, carácter común en especies que tienen reducidas las hojas portadoras de esporangios).
La prefoliación es circinada.
El carácter morfológico plesiomórfico (que ya existía en su antecesor común y todavía se mantiene en los descendientes) más característico de las polypodiopsidas es probablemente el esporangio, que es un leptosporangio, con todas sus variaciones.
El leptosporangio se caracteriza por emerger de un pie formado por 4-6 filas de células, que lo une a la hoja.
En general los esporangios están agrupados en "soros" en la cara abaxial de la hoja, o también puede encontrarse en porciones especializadas de las hojas (que entonces son "trofoesporofilos sectoriales"), o también pueden encontrarse en hojas fértiles especializadas en portar esporangios ("esporofilos"); menos común es encontrarlos dispersos por toda la cara abaxial de la hoja sin formar soros (condición "acrosticoide"), o dentro de esporocarpos (en Salviniales, adaptados a la vida acuática).
El leptosporangio usualmente posee un anillo de dehiscencia, que es un grupo de células de la pared del esporangio que se diferencian del resto de las células de la pared porque están más engrosadas en todas sus caras salvo la cara externa, y normalmente rodean al esporangio formando un anillo completo o incompleto (salvo en Osmundáceas, donde las células del "anillo" se agrupan en forma lateral, a veces llamado por eso "escudo de dehiscencia"). El anillo cumple la función de dehiscencia, ya que cuando se secan las células que lo forman, hacen presión en las paredes celulares para adaptarse al nuevo tamaño, pero como solo la pared externa es delgada, solo esa pared responde a la presión. Como resultado el anillo hace presión sobre toda la pared del esporangio hacia afuera, y la termina rompiendo debido a eso, el rompimiento brusco de la pared del esporangio catapulta a las esporas hacia afuera, y así se produce la dehiscencia.
La pared de los leptosporangios es de una sola célula de espesor a la madurez (salvo en Osmundáceas).
Casi todas las polypodiopsidas son homospóricas (un solo morfo de esporas), salvo Salviniales que desarrollaron la heterosporía (tienen dos morfos de esporas, esporas grandes o megasporas que dan gametos femeninos, y esporas pequeñas o microsporas que dan gametos masculinos).
Las esporas en general no son verdes.
Los gametófitos en general son exospóricos (de vida libre, desarrollados fuera de la espora) salvo en Salviniales, donde son endospóricos (se desarrollan hasta adultos dentro de la pared de la megaspora y la microspora, carácter usualmente asociado con la heterosporía).
Cuando son exospóricos, los gametófitos suelen ser verdes, fotosintéticos, y crecen en la superficie terrestre.
Luego de formado el cigoto, la primera división es vertical.
Véase también
[editar]- Clasificación de los organismos vegetales
- Para ver una lista completa de los taxones ver Sistema de clasificación de monilophytas de Smith 2006
- Para una descripción básica de morfología y anatomía de los helechos ver Pteridophyta
Enlaces externos
[editar]- Pryer's lab con los pdfs de las publicaciones en las que interviene la autora.
- Stevens, P. F. (2001 en adelante). Angiosperm Phylogeny Website. Actualizado regularmente.
Wikimedia Commons alberga una categoría multimedia sobre Polypodiidae.
Referencias
[editar]Libros
[editar]- Judd, W. S. Campbell, C. S. Kellogg, E. A. Stevens, P.F. Donoghue, M. J. 2002. Plant systematics: a phylogenetic approach, Second Edition. Sinauer Axxoc, USA.
Publicaciones
[editar]- Pryer, Kathleen M., Harald Schneider, Alan R. Smith, Raymond Cranfill, Paul G. Wolf, Jeffrey S. Hunt y Sedonia D. Sipes. 2001. "Horsetails and ferns are a monophyletic group and the closest living relatives to seed plants". Nature 409: 618-622 (pdf aquí).
- Pryer, Kathleen M., Eric Schuettpelz, Paul G. Wolf, Harald Schneider, Alan R. Smith y Raymond Cranfill. 2004. "Phylogeny and evolution of ferns (monilophytes) with a focus on the early leptosporangiate divergences". American Journal of Botany 91:1582-1598 (pdf aquí).
- A. R. Smith, K. M. Pryer, E. Schuettpelz, P. Korall, H. Schneider, P. G. Wolf. 2006. "A classification for extant ferns". Taxon 55(3), 705-731 (pdf aquí)
- Schneider, H., E. Schuettpelz, K.M. Pryer, R. Cranfill, S. Magallón, R. Lupia. 2004. "Ferns diversified in the shadow of angiosperms." Nature 428: 553-557. (pdf aquí)
Referencias citadas
[editar]- ↑ a b c Christenhusz et al. 2011. A linear sequence of extant families and genera of lycophytes and ferns. Phytotaxa 19: 7-54. (pdf)
- ↑ a b c Preface to “Linear sequence, classification, synonymy, and bibliography of vascular plants: Lycophytes, ferns, gymnosperms and angiosperms” pdf
- ↑ a b c Corrections to Phytotaxa 19: Linear sequence of lycophytes and ferns pdf
- ↑ a b c A. R. Smith, K. M. Pryer, E. Schuettpelz, P. Korall, H. Schneider, P. G. Wolf. 2006. "A classification for extant ferns". Taxon 55(3), 705-731 ( pdf)
- ↑ a b c Smith, A.R., Pryer, K.M., Schuettpelz, E., Korall, P., Schneider, H., & Wolf, P.G. (2008) Fern classification, pp. 417–467 en: Ranker, T.A., & Haufler, C.H. (eds.), Biology and Evolution of Ferns and Lycophytes. Cambridge , Cambridge University Press.
- ↑ Theodor Cole & Hartmut Hilger 2013 Trachaeophyte Phylogeny Archivado el 4 de marzo de 2016 en Wayback Machine.
| Control de autoridades |
|
|---|
Datos: Q3395756