Dinornithidae
| Moas | ||
|---|---|---|
| Rango temporal: Mioceno - Holoceno, 17 Ma - 0,0006 Ma | ||
 Moa gigante atacada por seres humanos (reconstrucción). | ||
| Estado de conservación | ||
|
Extinto desde ca. 1500 | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Aves | |
| Subclase: | Neornithes | |
| Superorden: | Palaeognathae | |
| Orden: |
Dinornithiformes Bonaparte, 1853 | |
| Géneros | ||
Los dinornitiformes o moas (Dinornithiformes) son un orden extinto de aves paleognatas.[1] Eran aves no voladoras que habitaban en Nueva Zelanda. Se conocen diez especies de diferentes tamaños, desde la pequeña Euryapteryx curtus, del tamaño de un gallo, hasta las moas gigantes (Dinornis), que medían cerca de tres metros de altura y tenían un peso de 250 kg. Su extinción se produjo hace quinientos años, causada por el ser humano.
Generalidades
[editar]
Los moas aparecieron en el Cretáceo hace más de noventa millones de años, siendo sus parientes vivos más próximos, los tinamúes de Surámerica, los kiwis, los casuarios y los emúes de Australia están más lejanamente emparentados siendo grupos divergentes de un mismo origen. Hasta tiempos modernos Nueva Zelanda y las islas que la rodean no estaban habitadas por mamíferos, por lo que era un ambiente que permitía a algunas aves adaptarse a hacer nidos en el suelo y a otras volverse no voladoras. Los moas se extinguieron alrededor del año 1400, poco después de que los primeros cazadores maoríes llegaran a las islas. Algunos creen que existieron pequeños grupos de moas hasta fines del siglo XVIII o inicios del XIX. De hecho, se cuenta algún avistamiento por parte de marineros a las órdenes de James Cook, y llama la atención el estado de conservación de algunas plumas de estas aves. Sin embargo no hay pruebas físicas.
La filogeografía de este grupo está bastante bien establecida y es un ejemplo de la acción de varios mecanismos de especiación. Alrededor de ochenta y dos millones de años atrás, los ancestros de este grupo quedaron aislados de las restantes aves rátidas, cuando Nueva Zelanda se separó de Gondwana, resultando en la separación física de los grupos. Este mecanismo es llamado especiación alopátrica. Pasado un tiempo, los ancestros de los géneros sobrevivientes, se adaptaron a diferentes nichos ecológicos. Esto dio lugar a aislamiento reproductivo, y es ejemplo de especiación ecológica.
En el Plioceno, alrededor de cinco millones de años atrás, la formación de los Alpes del Sur diversificó el paisaje y proveyó nuevas oportunidades para la especiación dentro de los nuevos géneros. Unos tres millones de años atrás, varios linajes se adaptaron a altitudes altas y bajas. El linaje de bajas temperaturas (montaña, altas altitudes, edades glaciales) originó especies más resistentes a la baja temperatura y de menor tamaño, mientras que el de bajas altitudes o clima más suave, dio lugar a las distintas especies más extendidas.
Con el aislamiento de unas pocas aves errantes que llegan a una isla adecuada, surge rápidamente una especie isleña por divergencia con las poblaciones de la especie de los territorios de origen. Tanto las varias especies de kiwis como los moas son el resultado de la migración de un número limitado de individuos a islas y la adaptación posterior al hábitat de esas islas.
Finalmente, en tiempos recientes, poblaciones de aves del territorio neozelandés quedaron divididas entre la Isla Norte y la Isla Sur debido al ascenso del nivel del mar cuando los glaciares continentales se derritieron al final del Pleistoceno.
Antes del arribo de los maoríes el equilibrio de los moas se había saldado con un descenso en su densidad en el último millón de años, causado por varios factores: los periodos glaciales, en los que casi toda la Isla Sur fue aislada en pequeños territorios por el hielo; el cambio climático que disminuyó los espacios cubiertos de tussok, sustituidos por selvas y bosques densos debido al aumento gradual de la temperatura y la humedad; la diversificación en especies para evitar la competencia ecológica, la predación de las águilas de Haast (Harpagornis moorei), la mayor ave de presa conocida, hoy también extinta.
Los moas, a diferencia de otras ratites, como el avestruz, el emú, el casuario o el ñandú, durante la evolución habían perdido completamente las alas, tal como ocurre con los supervivientes kiwis, que se consideraban emparentados con los moas.
La fauna de Nueva Zelanda evolucionó por largo tiempo en ausencia de humanos y otros mamíferos. Sólo unas pocas especies de murciélagos y mamíferos marinos estaban presentes antes de la colonización por los humanos, y los únicos depredadores eran aves de presa que cazaban usando la vista. Estas circunstancias influenciaron la evolución de las aves de Nueva Zelanda, por ejemplo, las adaptaciones a la falta de vuelo del kakapo y la anidación en el suelo del kea. Los polinesios llegaron a las islas entre el año 800 y 1300 de nuestra era,[2] e introdujeron el perro polinesio (kuri) en las islas.
Esto fue desastroso para la fauna nativa, porque los predadores mamíferos pueden localizar sus presas por el olor, y la fauna nativa no había evolucionado para defenderse de ellos. Los principales depredadores eran aves: águilas (aguiluchos de Eyles, Kāhu y águila de Haast ), halcones (Kārearea) y búhos (Whēkau y Ruru). Muchas de las adaptaciones de las aves neozelandesas refleja el contexto único en el que evolucionaron. Este balance único fue perturbado con la llegada de los Polinesios, quienes introdujeron la rata polinesia y el Kuri (perro polinesio) en las islas. Posteriormente, los europeos introdujeron muchas más especies, incluyendo grandes herbívoros y predadores mamíferos.
Para evitar la sobrepoblación, los moas alcanzaban muy tardíamente su madurez reproductiva: a los diez años de nacidos, como la especie de moa gigante Dinornis robustus. Este largo tiempo para su madurez facilitó su extinción, ya que no se dio una suficiente tasa de reposición ante el ritmo con el que eran cazados por los maoríes. Los moas se alimentaban de semillas, frutas, hierba e incluso ramas. Para referirse a estas aves los maoríes usan tanto en singular como en plural la palabra moa sin ese final.
Dimorfismo y taxonomía
[editar]
Habitantes de Nueva Zelanda, las moas son toda una familia de aves de la que se conocen once especies de distintos tamaños, en seis géneros, yendo desde el tamaño de un gallo a otras midiendo hasta tres metros de altura y pesando doscientos cincuenta kilogramos.
Hasta fechas recientes existía una clasificación que dividía en dos especies a las hembras y machos de cada especie de moa. Esto era debido al notorio dimorfismo sexual, ya que las hembras de muchas especies eran aproximadamente una vez y media más altas y corpulentas que los machos. Sin embargo, gracias a los análisis de ADN se ha podido elucidar la razón de que en algunas especies sólo se encontrasen ejemplares de uno de los sexos y no de ambos.
Actualmente se sabe que Euryapteryx curtus era concretamente la hembra del Euryapteryx exilis; lo mismo ocurría con las siguientes parejas: Emeus hutonii / Emeus crassus, Pachyornis septentrionalis / Pachyornis mappini, Dinornis giganteus / Dinornis robustus y Dinornis novaezelandiae / Dinornis struthoides. Por esta doble clasificación, en la actualidad la taxonomía utiliza, generalmente, para designar a estas especies los nombres aplicados a las mucho más corpulentas hembras.
Taxonomía
[editar]El orden Dinornithiformes comprende:[3]
- Orden †Dinornithiformes - moa
- Familia Dinornithidae - Moa gigante
- Género Dinornis (moas gigantes)
- Moa gigante de la isla Norte (Dinornis novaezealandiae) (Isla Norte, Nueva Zelanda)
- Moa gigante de la isla Sur (Dinornis robustus) (Isla Sur, Nueva Zelanda)
- Género Dinornis (moas gigantes)
- Familia Dinornithidae - Moa gigante
- Familia Emeidae - Moa Menor
- Género Anomalopteryx
- Moa de los arbustos, Anomalopteryx didiformis (Isla Sur, Nueva Zelanda)
- Género Emeus
- Moa oriental, Emeus crassus (Isla Sur, Nueva Zelanda)
- Género Euryapteryx
- Moa costero, Euryapteryx curtus (Isla Norte, Nueva Zelanda)
- Género Pachyornis
- Moa pesado, Pachyornis elephantopus (Isla Sur, Nueva Zelanda)
- Moa de patas fornidas, Pachyornis geranoides (Isla Sur, Nueva Zelanda)
- Moa crestado, Pachyornis australis (Isla Sur, Nueva Zelanda)[4]
- Género Anomalopteryx
- Familia Megalapterygidae
- Género Megalapteryx
- Moa de tierraarriba, Megalapteryx didinus (Isla Sur, Nueva Zelanda)
- Género Megalapteryx
- Familia Emeidae - Moa Menor
Relaciones evolutivas
[editar]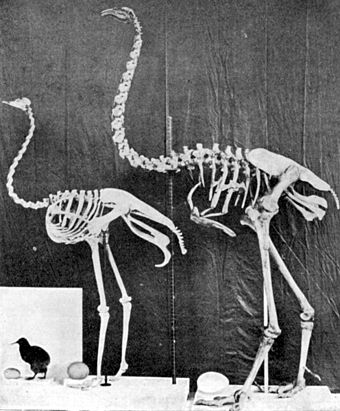
Los parientes más cercanos del moa son unas pequeñas aves terrestres sudamericanas llamadas tinamús, que pueden volar.[5][6][7][8] Anteriormente, se pensaba que el kiwi, el emú australiano y el casuario[9] eran los parientes más cercanos del moa.
Aunque a finales del siglo XIX y principios del XX se describieron docenas de especies, muchas se basaban en esqueletos parciales y resultaron ser sinónimos. En la actualidad, se reconocen formalmente once especies, aunque estudios recientes realizados con ADN antiguo recuperado de huesos de colecciones de museos sugieren la existencia de linajes distintos dentro de algunas de ellas. Un factor que ha causado mucha confusión en la taxonomía de los moa es la variación intraespecífica del tamaño de los huesos, entre periodos glaciares e interglaciares (véase la regla de Bergmann y la regla de Allen), así como el dimorfismo sexual evidente en varias especies. Dinornis parece haber tenido el dimorfismo sexual más pronunciado, con las hembras siendo hasta un 150% más altas y un 280% más pesadas que los machos, tanto más grandes que fueron clasificados como especies separadas hasta 2003.[10][11] Un estudio de 2009 mostró que Euryapteryx curtus y E. gravis eran sinónimos.[12]
Un estudio de 2010 explicó las diferencias de tamaño entre ellos como dimorfismo sexual.[13] En cambio, un estudio morfológico de 2012 los interpretó como subespecies.[14]
Los análisis de ADN antiguo han determinado la existencia de una serie de linajes evolutivos crípticos en varios géneros de moa,[15] que con el tiempo podrían clasificarse como especies o subespecies; Megalapteryx benhami (Archey) se sinonimiza con M. didinus (Owen) porque los huesos de ambos comparten todos los caracteres esenciales. Las diferencias de tamaño pueden explicarse por una clina norte-sur combinada con una variación temporal tal que los especímenes eran más grandes durante el periodo glaciar de Otiran (la última era glaciar en Nueva Zelanda). Se conoce una variación temporal similar en el tamaño del Pachyornis mappini de la Isla Norte.[16] Algunas de las demás variaciones de tamaño de las especies de moa pueden explicarse probablemente por factores geográficos y temporales similares.[17]
Los restos más antiguos de moa proceden de la fauna del Mioceno de Saint Bathans. Conocidos a partir de múltiples cáscaras de huevo y elementos de las extremidades posteriores, representan al menos dos especies ya de tamaño bastante grande.[18]
Referencias
[editar]- ↑ Livezey, Bradley C. & Zusi, Richard L. (2007): Higher-order phylogeny of modern birds (Theropoda, Aves: Neornithes) based on comparative anatomy. II. Analysis and discussion. Zoological Journal of the Linnean Society 149 (1), 1–95.
- ↑ Douglas G. Sutton, ed. (1994). The Origins of the First New Zealanders. Auckland: Auckland University Press. ISBN 1869400984.
- ↑ Davies, S. J. J. F. (2003). «Moas». En Hutchins, Michael, ed. Grzimek's Animal Life Encyclopedia. Volumen 8: Birds I: Tinamous and Ratites to Hoatzins (en inglés) (2ª edición). Detroit: Gale. pp. 95-98. ISBN 0-7876-5784-0.
- ↑ Stephenson, Brent (1 de mayo de 2009). «New Zealand Recognised Bird Names (NZRBN) database». Nueva Zelanda: Ornithological Society of New Zealand. Archivado desde el original el 25 de abril de 2015. Consultado el 5 de octubre de 2010.
- ↑ Phillips, et al. (2010)
- ↑ Allentoft, M.E.; Rawlence, N.J. (20 de enero de 2012). «Moa's Ark or volant ghosts of Gondwana? Insights from nineteen years of ancient DNA research on the extinct moa (Aves: Dinornithiformes) of New Zealand». Annals of Anatomy - Anatomischer Anzeiger 194 (1): 36-51. PMID 21596537. doi:10.1016/j.aanat.2011.04.002.
- ↑ Mitchell, K.J.; Llamas, B.; Soubrier, J.; Rawlence, N.J.; Worthy, Trevor; Wood, J.; Lee, M.S.Y.; Cooper, A. (23 de mayo de 2014). «Ancient DNA reveals elephant birds and kiwi are sister taxa and clarifies ratite bird evolution». Science (en inglés) 344 (6186): 898-900. Bibcode:2014Sci...344..898M. PMID 24855267. S2CID 206555952. doi:10.1126/science.1251981. hdl:2328/35953. Archivado desde el original el 30 de mayo de 2019.
- ↑ Baker, A.J.; Haddrath, O.; McPherson, J.D.; Cloutier, A. (2014). «Genomic Support for a Moa-Tinamou Clade and Adaptive Morphological Convergence in Flightless Ratites». Molecular Biology and Evolution 31 (7): 1686-1696. PMID 24825849. doi:10.1093/molbev/msu153.
- ↑ Turvey et al. (2005)
- ↑ Huynen, L.J., et al. (2003)
- ↑ Bunce, M., et al. (2003)
- ↑ Bunce, M.; Worthy, Trevor; Phillips, M.J.; Holdaway, Richard; Willerslev, E.; Haile, J.; Shapiro, B.; Scofield, R.P.; Drummond, A.; Kamp, P.J.J.; Cooper, A. (2009). «The evolutionary history of the extinct ratite moa and New Zealand Neogene paleogeography». Proceedings of the National Academy of Sciences 106 (49): 20646-20651. Bibcode:2009PNAS..10620646B. PMC 2791642. PMID 19923428. doi:10.1073/pnas.0906660106.
- ↑ Gill, B.J. (2010). «Regional comparisons of the thickness of moa eggshell fragments (Aves: Dinornithiformes). In Proceedings of the VII International Meeting of the Society of Avian Paleontology and Evolution, ed. W.E. Boles and Trevor Worthy». Records of the Australian Museum (en inglés) 62: 115-122. doi:10.3853/j.0067-1975.62.2010.1535. Archivado desde el original el 11 de abril de 2019.
- ↑ Worthy, Trevor; Scofield, R.P. (2012). «Twenty-first century advances in knowledge of the biology of moa (Aves: Dinornithiformes): A new morphological analysis and moa diagnoses revised». New Zealand Journal of Zoology 39 (2): 87-153. S2CID 83768608. doi:10.1080/03014223.2012.665060.
- ↑ Baker, A.J.; Huynen, L.J.; Haddrath, O.; Millar, C.D.; Lambert, D.M. (2005). «Reconstructing the tempo and mode of evolution in an extinct clade of birds with ancient DNA: The giant moas of New Zealand». Proceedings of the National Academy of Sciences 102 (23): 8257-8262. Bibcode:2005PNAS..102.8257B. PMC 1149408. PMID 15928096. doi:10.1073/pnas.0409435102.
- ↑ Worthy (1987)
- ↑ Worthy, et al. (1988)
- ↑ Tennyson, A.J.D.; Worthy, Trevor; Jones, C.M.; Scofield, R.P.; Hand, S.J. (2010). «Moa's Ark: Miocene fossils reveal the great antiquity of moa (Aves: Dinornithiformes) in Zealandia». Records of the Australian Museum (en inglés) 62: 105-114. doi:10.3853/j.0067-1975.62.2010.1546. Archivado desde el original el 11 de abril de 2019.
Véase también
[editar]Enlaces externos
[editar]Wikimedia Commons alberga una galería multimedia sobre Dinornithidae.
- Moa Central (en inglés)
Datos: Q452969
Especies: Dinornithiformes