Sac vitel·lí
| Aquest article o secció no cita les fonts o necessita més referències per a la seva verificabilitat. |
El sac vitel·lí [1] o coberta vitel·lina[2] és un annex membranós adossat a l'embrió que proveeix nutrients i oxigen a l'embrió en peixos, taurons, rèptils, aus i mamífers primitius, alhora que elimina deixalles metabòliques. En l'embrió humà funciona com a sistema circulatori en les etapes de desenvolupament, abans que comenci la circulació interna.
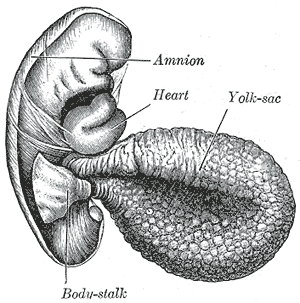
El sac vitel·lí està cobert per una prima capa mucosa i una paret més interna rica en vasos sanguinis que seran aportats al comunicat embrionari. Després de la gastrulació, el sac vitel·lí queda suspès i sostingut del cos embrionari, de manera que dona forma al conducte vitel·lí.
Mitjançant el desenvolupament fetal, i principalment a partir de la 9a o 10a setmana de desenvolupament, el sac vitel·lí comença a degenerar de manera natural per diverses causes, com la seva compressió causa de l'augment de l'espai ocupat per l'amni. Durant el període primerenc del desenvolupament embrionari, el sac vitel·lí és l'estructura de major dimensió, amb una forma ovalada, però que amb el desenvolupament, adquireix una aparença piriforme.[3]
No es coneix quant temps roman actiu el sac vitel·lí.
Estructura del sac vitel·lí
modificaEl sac vitel·lí està constituït morfològicament per 3 capes, que són: capa endodèrmica, capa mesenquimàtica i capa mesotelial:
- a) Capa endotèrmica:
glucogen. Aquesta monocapa després es tornarà dinàmica i funcionalment essencial per l'embrió. Es formen després una sèrie de cordons endodèrmics que penetren a la capa mesenquimàtica. Entre aquests cordons es creen petits orificis (vesícules endodèrmiques) que contenen substància d'excreció produïda per cèl·lules circumdants. Aquestes vesícules amb el temps tendiran a fusionar fins a generar grans vesícules que es relacionen amb la xarxa de vasos sanguinis presents en la mesenquimàtica circumdant. Aquesta capa presenta una degeneració funcional amb el desenvolupament de l'embrió.
- b) Capa mesenquimàtica:
- c) Capa mesotelial:
Està constituït per una monocapa de cèl·lules planes unides entre si. Confronta a l'espai intervitel·lí del sac. Les cèl·lules d'aquesta capa presenten una gran quantitat de microvellositats. A partir de la setmana setena, s'observen canvis degeneratius en l'epiteli, es formen rugositats profundes de la superfície i augment en la quantitat de mucus, la qual cosa provoca el col·lapse de les microvellositats. Aquestes pateixen un procés de fragmentació i es desprenen de la superfície cel·lular. Els orgànuls citoplasmàtics involucionen quedant el citoplasma molt vacuolat.
Vitel
modificaQuan ocorre la fecundació entre un gàmeta masculí (espermatozoide) i un altre femení (ovòcit), l'espermatozoide proveeix únicament material genètic, mentre que l'oòcit (més gran) aporta o sintetitza també les substàncies alimentàries necessàries per al desenvolupament embrionari del zigot. Aquest material nutritiu és denominat vitel i es constitueix de diferents substàncies proteiques, algunes sintetitzades en l'ovòcit i altres provinents del torrent sanguini i absorbides per l'oòcit. Un cop produït el vitel, aquest és transportat cap a l'hemisferi o pol vegetal de l'ou (meitat inferior de l'ou) i en general aquesta regió conté molt més vitel que l'hemisferi o pol animal (meitat superior de l'ou); pol animal o vegetal es deu als moviments observats en alguns embrions models (com el de l'ou d'un amfibi) on les cèl·lules derivades de la meitat superior estan activament mòbils (o animades) i les de la meitat inferior són gairebé immòbils (o estàtiques com les plantes). Mentre que les plaquetes del vitel s'acumulen més densament al pol vegetal de l'ovòcit, els grànuls de glucogen, els ribosomes, les vesícules lipídiques i el reticle endoplasmàtic es troben cap al pol animal. La quantitat de vitel dependrà del tipus de desenvolupament, és a dir, entre més vitel s'acumuli, més durarà el procés embrionari i la segmentació prengués un camí diferent. La segmentació de l'ovòcit comença en el pol animal i descendeix lentament cap al pol vegetal, i segons la concentració de vitel, es produirà una segmentació holoblàstica o meroblàstica. En moltes espècies, l'oòcit (gàmeta femení en desenvolupament) és flagel·lat inicialment fins a l'ovogènesi on es perd el flagel. Aquesta característica es pot veure com un canvi secundari propi d'aquest gàmeta femení ia més marca on serà el futur pol animal (Gilbert, 2005).
Patrons principals de segmentació (segons la quantitat i distribució del vitel)
modificaSegmentació holoblàstica
modifica- Isolecítica: Poc vitel i distribuït equitativament per tot l'ou produint segmentació radial (equinoderms i anfioxus), espiral (anèl·lids, mol·luscs i cucs plans), bilateral (tunicats), i rotacional (mamífers i nematodes).
- Mesolecítica: Quantitat de vitel moderada que produeix segmentació radial desplaçada (amfibis).
Segmentació meroblàstica
modifica- Telolecítica: vitel dens al llarg de la major part de l'ou i més encara sobre el pol vegetal. S'observa segmentació bilateral (mol·luscs cefalòpodes), i discoïdal (peixos, rèptils i aus).
- Centrolecítica: vitel al centre de l'ou amb segmentació superficial (present en la majoria d'insectes).
Producció de vitel (vitel·logènesi)
modificaLa vitel·logènesi es produeix quan l'oòcit arriba l'estadi de diplot de la profase meiòtica. El vitel no és una sola subunitat o compost, sinó una barreja de materials proteics utilitzats per a la nutrició embrionària. Algunes proteïnes principals del vitel són sintetitzades en l'ovòcit (com en el cas d'alguns anèl·lids i crancs de riu), però altres també molt importants són portades a l'ovòcits pel torrent sanguini.
Quan la femella està fèrtil, la seva glàndula hipòfisi secreta hormones que li indiquen l'ovari de produir estrogen. L'estrogen és una hormona que pot instruir el fetge a produir i secretar proteïnes vitel·lines com ara la vitel·logenina, que són després transportades a través de la sang cap als ous en fase de creixement (abans de l'estimulació per estrogen no es veu cap ARNm de vitel·logenina a les cèl·lules hepàtiques, el qual és el primer pas per a la inducció de la síntesi d'aquesta proteïna. Després de l'estímul es poden observar unes 50.000 molècules de ARNm de vitel·logenina a les cèl·lules hepàtiques). Aquesta substància (la vitel·logenina) i les altres proteïna vitel·linas que no són sintetitzades en l'ovòcit, són enviada a l'ou en creixement a través de la sang on l'oòcit l'absorbeix i l'emmagatzema.
Referències
modifica- ↑ «Sac vitel·lí». Gran Enciclopèdia Catalana. Barcelona: Grup Enciclopèdia Catalana.
- ↑ «Sac vitel·lí». Cercaterm. TERMCAT, Centre de Terminologia.
- ↑ Guillermo Palomero. Lecciones de Embriología. Universidad de Oviedo, 2000, p. 420–. ISBN 978-84-8317-079-3.
Bibliografia
modifica- Gilbert, S. F., Biologia del Desenvolupament. 2005
- Wilt FH, Hake S., Principles of Developmental Biology. 2004