Proteïna
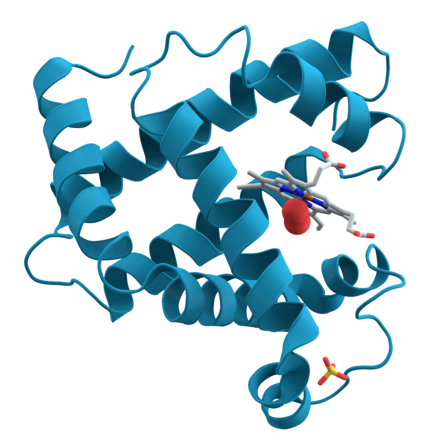
Les proteïnes, també denominades polipèptids, són compostos orgànics fets d'aminoàcids arranjats en una cadena lineal i units per enllaços peptídics entre els grups carboxil i amino de residus adjacents. La seqüència d'aminoàcids d'una proteïna és definida per la seqüència d'un gen, que està codificada al codi genètic.[1] En general, el codi genètic especifica vint aminoàcids estàndard, però en alguns organismes el codi genètic pot incloure la selenocisteïna i —en certs arqueus— pirrolisina. Poc després o fins i tot durant la síntesi, els residus d'una proteïna sovint són modificats químicament per la modificació posttraduccional, que altera les propietats físiques i químiques, el plegament, l'estabilitat, l'activitat i, en última instància, la funció de les proteïnes. Les proteïnes també poden col·laborar per complir una funció determinada, i sovint s'associen per formar complexos estables.[2] Les proteïnes fan moltes funcions, les més importants són la funció metabòlica i també la funció de transport d'oxigen i d'anticossos.
Com altres macromolècules biològiques com els polisacàrids i els àcids nucleics, les proteïnes són parts essencials dels organismes i participen en la pràctica totalitat dels processos cel·lulars. Moltes proteïnes són enzims que catalitzen reaccions bioquímiques i són essencials per al metabolisme. Les proteïnes també tenen funcions estructurals o mecàniques, com l'actina i la miosina dels músculs o les proteïnes del citoesquelet, que formen una carcassa que manté la forma de la cèl·lula. Altres proteïnes són importants en la senyalització cel·lular, la resposta immunitària, l'adherència cel·lular i el cicle cel·lular. Les proteïnes també són necessàries en la dieta dels animals, car no poden sintetitzar tots els aminoàcids que necessiten i han d'obtenir aminoàcids essencials del menjar. Mitjançant el procés de la digestió, els animals descomponen les proteïnes que han ingerit en aminoàcids lliures que posteriorment són utilitzats en el metabolisme.
Les proteïnes foren descrites i anomenades per primer cop pel químic suec Jöns Jacob Berzelius el 1838. Tanmateix, el paper essencial de les proteïnes en els éssers vius no fou apreciat completament fins al 1926, quan James Batcheller Sumner demostrà que l'enzim ureasa era una proteïna.[3] La primera proteïna que fou seqüenciada fou la insulina, per Frederick Sanger, que guanyà el Premi Nobel pel seu treball el 1958. Les primeres estructures proteiques en ser determinades foren la de l'hemoglobina i la de la mioglobina, per Max Perutz i Sir John Cowdery Kendrew, respectivament, el 1958.[4][5] L'estructura tridimensional d'ambdues proteïnes fou determinada per primer cop per anàlisi de difracció de rajos X; Perutz i Kendrew compartiren el Premi Nobel de Química del 1962 per aquests descobriments. Les proteïnes es poden purificar de la resta de components cel·lulars mitjançant una varietat de tècniques com ara ultracentrifugació, precipitació, electroforesi i cromatografia; l'arribada de l'enginyeria genètica ha fet possible una sèrie de mètodes per facilitat la purificació. Els mètodes utilitzats habitualment per estudiar l'estructura i el funcionament de les proteïnes inclouen la immunohistoquímica, la mutagènesi dirigida i l'espectrometria de masses.
Bioquímica
[modifica]
Les proteïnes són polímers lineals formades per sèries de fins a vint L-α-aminoàcids diferents. Tots els aminoàcids presenten trets estructurals comuns, incloent-hi un α-carboni al qual estan units un grup amino, un grup carboxil i una cadena lateral variable. La prolina és l'única que difereix d'aquesta estructura bàsica, car conté un anell inusual al grup amino de l'extrem-N, que força el fragment d'amida CO–NH en una conformació fixa.[6] Les cadenes laterals dels aminoàcids estàndard, detallats a la llista d'aminoàcids estàndard, tenen una gran varietat d'estructures i propietats químiques; és l'efecte combinat de totes les cadenes laterals dels aminoàcids d'una proteïna el que determina en última instància la seva estructura tridimensional i la seva reactivitat química.[7]


Els aminoàcids d'una cadena polipeptídica estan units per enllaços peptídics. Una vegada enllaçats a la cadena proteica, un aminoàcid individual és anomenat "residu", mentre que les sèries unides d'àtoms de carboni, nitrogen i oxigen són coneguts com a "cadena principal" o "tronc proteic".[8] L'enllaç peptídic té dues formes de ressonància que contribueixen un cert caràcter de doble enllaç i inhibeixen la rotació al voltant del seu eix, de manera que els alfa-carbonis són més o menys coplanars. Els altres dos angles dièdrics de l'enllaç peptídic determinen la forma local que assumeix el tronc proteic.[9] L'extrem de la proteïna amb el grup carboxil lliure és conegut com a carboxil terminal, mentre que l'extrem amb el grup amino lliure és l'amino terminal.
Els mots "proteïna", "polipèptid" i "pèptid" són una mica ambigus i el seu significat es pot solapar. "Proteïna" se sol utilitzar per referir-se a la molècula biològica completa en una conformació estable, mentre que "pèptid" està reservat generalment per oligòmers curts d'aminoàcids, que sovint manquen d'estructura tridimensional estable. Tanmateix, el límit entre els dos no està ben definit i sovint es marca a prop de 20-30 residus."[10] Polipèptid" es pot referir a qualsevol cadena lineal d'aminoàcids, sovint sense prendre en compte la seva llargada, però sovint implica l'absència d'una conformació definida.

Estructura i plegament
[modifica]Molts dels enllaços que formen una cadena polipeptídica permeten la lliure rotació dels àtoms que uneixen, la qual cosa confereix a l'esquelet de la proteïna una gran flexibilitat. Per tant, qualsevol molècula proteica pot, en principi, adoptar un nombre il·limitat de formes diferents (conformacions). No obstant això, la majoria de les cadenes polipeptídiques es pleguen adoptant una sola conformació particular. Això és a causa del fet que els esquelets dels diferents aminoàcids, a manera de cadenes laterals de la cadena principal, s'associen entre si i amb l'aigua formant diversos enllaços no covalents febles. Sempre que les cadenes laterals apropiades es trobin en posicions crucials en la cadena, es desenvoluparan importants forces que fan que una conformació particular sigui especialment estable.
Un dels factors més importants que condicionen el plegament d'una proteïna és la distribució de les seves cadenes laterals polars i no polars. Les cadenes laterals hidròfobes, molt abundants, tendeixen a agrupar-se en l'interior de la molècula, la qual cosa els permet evitar el contacte amb l'entorn aquós. Per contra, les cadenes laterals polars tendeixen a disposar-se prop de la part externa de la molècula proteica, on poden interaccionar amb l'aigua i altres molècules polars. Al descriure l'estructura d'una proteïna, resulta útil distingir diversos quatre nivells d'organització, cadascun dels quals es construeix sobre el nivell anterior d'una forma jeràrquica.
Estructura primària
[modifica]És comuna a totes les proteïnes i és la seqüència lineal d'aminoàcids que la integren; és a dir, indica els aminoàcids que la formen i l'orde en què es troben units. És l'estructura més senzilla i, tanmateix, la més important, ja que determina la resta de les estructures proteiques amb nivells superiors d'organització.
Una característica d'aquesta estructura és la seva disposició en ziga-zaga, deguda al caràcter planar de l'enllaç peptídic, cosa que provoca la rotació dels aminoàcids sobre els Cα a fi d'equilibrar les forces d'atracció que es puguin generar. Amb els vint aminoàcids possibles, el nombre de polipèptids que es poden formar és de 20n, en què n és el nombre d'aminoàcids presents en la cadena. Com que la majoria de pèptids contenen més de cent aminoàcids, fins i tot milers, la varietat de possibles seqüències és pràcticament il·limitada.
Estructura secundària
[modifica]És la disposició espacial que adopta la seqüència d'aminoàcids o estructura primària a fi de ser estable, i és una conseqüència directa de la capacitat de gir que tenen els carbonis α dels aminoàcids. Els models més freqüents són l'α-hèlix i la conformació β o làmina plegada.
- Hèlix alfa o α-hèlix: en aquest tipus d'estructura, la cadena polipeptídica s'enrotlla en espiral sobre si mateixa a causa dels girs que es produeixen entorn de carboni α de cada aminoàcid. Aquesta estructura es manté gràcies als enllaços d'hidrogen intracatenaris formats entre el grup –NH de l'enllaç peptídic i el grup –C=O del quart aminoàcid que el segueix. Quan es forma l'α-hèlix, tots els grups –C=O queden orientats en la mateixa direcció, mentre que els –NH s'orienten en direcció contrària i els radicals dels aminoàcids queden dirigits cap a l'exterior de l'α-hèlix. La rotació té lloc cap a la dreta, de manera que cada aminoàcid gira 100° respecte a d'anterior. Això explica la presència de 3,6 residus aminoàcids per cada volta completa.
- Làmina β o conformació β o làmina [beta] plegada: a diferència de l'α-hèlix, aquí no interaccionen segments continus d'una única cadena polipeptídica, sinó diferents combinacions de seccions que no necessàriament van seguides una després de l'altra i que pertanyen a una o més cadenes polipetídiques. Els fragments de cadenes β estan ordenats un al costat de l'altre de manera que es formen ponts d'hidrogen entre els grups CO i NH de cadenes veïnes. Les cadenes que interaccionen poden ésser paral·leles (les cadenes polipeptídiques es disposen en el mateix sentit N-C) o antiparal·leles (les cadenes polipeptídiques s'alternen en les direccions N-C i C-N). En ambdós casos, els radicals dels aminoàcids s'orienten envers ambdues bandes de la fulla, de manera alterna. Ocasionalment es formen làmines plegades β mixtes amb orientació paral·lela a un costat de la cadena i antiparal·lela a l'altre. Empíricament es demostra que la isoleucina, la valina, la treonina, la fenilalanina i la tirosina afavoreixen la formació d'estructures de làmina plegada; això no obstant, és estrany trobar restes de prolina en les β làmina plegades.
- Hèlix de col·lagen: en les molècules de col·lagen, tres cadenes polipeptídiques distintes riques en l'aminoàcid prolina, estan enrotllades entre si generant una triple hèlix regular. Aquestes molècules de col·lagen estan, al seu torn, empaquetades formant fibril·les en les quals les molècules de col·lagen adjacents estan unides per enllaços covalents donant a les fibril·les una enorme resistència la tensió.
- Elastina: les cadenes polipeptídiques de l'elastina, relativament lliures i desestructurades, estan unides per enllaços covalents creuats, generant una trama elàstica semblant al cautxú que permet que teixits tals com els de les artèries i dels pulmons es deformin i dilatin sense sofrir cap dany. L'elasticitat és deguda a la capacitat de les diferents molècules proteiques per a desenrotllar-se de manera reversible cada vegada que se'ls apliqui una força de tensió.
Estructura terciària
[modifica]L'estructura terciària de les proteïnes, especialment de les que tenen un pes molecular elevat, està constituïda per diversos dominis o unitats compactes de 50 a 300 aminoàcids connectades a través de l'esquelet polipeptídic. Un domini proteic pot concebre's com la unitat estructural bàsica d'una estructura proteica. El nucli de cada domini està compost fonamentalment per un conjunt de làminesβ, α-hèlix o d'ambdues coses. Aquestes estructures secundàries regulars estan afavorides a causa del fet que permeten la formació d'una gran quantitat d'enllaços d'hidrogen entre els àtoms de l'esquelet de la proteïna, la qual cosa és essencial per a estabilitzar l'interior del domini, on l'aigua no és assequible per a forma enllaços d'hidrogen amb l'oxigen polar del carbonil o amb l'hidrogen de l'amida de l'enllaç peptídic. Existeix un nombre limitat de maneres de combinar α-hèlix amb làminesβ per a formar una estructura globular, pel que determinades combinacions d'aquests elements, denominats motius, es presenten repetidament en el nucli de moltes proteïnes no relacionades entre si. Diverses combinacions de motius formen el domini proteic, en el qual la cadena polipeptídica tendeix a corbar-se dalt i baix al llarg de tota l'estructura, de vegades formant una làmina β o una α-hèlix. Les característiques d'una proteïna i, del que és més important, les funcions biològiques que du a terme depenen de la seva estructura terciària.
Estructura quaternària
[modifica]Encara que moltes proteïnes estan formades per una única cadena polipeptídica, també n'hi ha moltes que estan constituïdes per més d'una subunitat o protòmer. L'estructura quaternària fa referència a aquesta associació de protómers per constituir la proteïna biològica activa. Els protòmers es poden unir dèbilment entre si mitjançant enllaços d'hidrogen o forces de Van der Waals, i de vegades, tot i que no és habitual, aquesta unió es pot establir mitjançant ponts disulfur.
Biosíntesi
[modifica]
Les proteïnes es munten a partir d'aminoàcids usant la informació codificada als gens. Cada proteïna té la seva pròpia seqüència d'aminoàcids especificada per la seqüència de nucleòtids del gen que codifica cada proteïna. El codi genètic és un joc de codons format per tres nucleòtids i cada combinació de tres nucleòtids codifica un aminoàcid, per exemple AUG codifica la metionina. Pel fet que l'ADN conté quatre nucleòtids, el nombre total de codons possibles és de 64; així doncs, hi ha redundància en el codi genètic: alguns aminoàcids venen codificats per més d'un codó. Els gens codificats en l'ADN es transcriuen en pre-ARNm per proteïnes com l'ARN polimerasa. La majoria d'organismes que processen el pre-mRNA (també conegut com a transcrit primari) usant diverses formes de modificació post-transcripcional per a la maduració de l'ARNm, el qual s'usa llavors com a motlle per a la síntesi proteica al ribosoma. En procariotes l'ARNm també pot usar-se tan aviat com es produeix o unir-se a un ribosoma després d'haver-se separat del nucleoide. En contrast, els eucariotes sintetitzen l'ARNm al nucli cel·lular i llavors travessen la membrana nuclear fin el citoplasma, on llavors té lloc la síntesi proteica. La ràtio de síntesi de proteïnes és més gran en procariotes que en eucariotes, assolint 20 aminoàcids per segon.[11]
El procés de síntesi de proteïnes a partir d'un motlle d'ARNm es coneix com a traducció. L'ARNm és carregat a l'interior del ribosoma i es llegeixen tres nucleòtids alhora en correspondència de cada codó al seu parell de bases de l'anticodó localitzat a l'ARN de transferència (ARNt), el qual carrega l'aminoàcid corresponent al codó i reconeix. L'enzim aminoacil ARNt sintetasa «carrega» les molècules d'ARNt] amb l'aminoàcid específic. El polipèptid en elongació sovint s'anomena «cadena naixent». Les proteïnes sempre són biosintetitzades de l'extrem N terminal al C terminal.
La mida d'una proteïna es pot mesurar pel nombre d'aminoàcids que conté i per la seva massa molecular total la que és normalment mesurada en daltons (sinònim d'unitat de massa atòmica) i el múltiple, el kilodalton (kDa) més usat. Les proteïnes dels llevats tenen una mitjana de 466 aminoàcids de longitud i uns 53 kDa de massa.
Les proteïnes més grans conegudes són les titines, un component del sarcòmer del múscul amb una massa molecular d'almenys 3.000 kDa i una longitud total d'almenys 27.000 aminoàcids.[12]
Síntesi química
[modifica]Les proteïnes petites també es poden sintetitzar químicament pel conjunt de mètodes coneguts com a síntesi de pèptids, els quals es relacionen amb tècniques de síntesi orgànica com ara el lligament químic per a produir pèptids en multitud de camps.[13] La síntesi química permet la introducció d'aminoàcids no naturals a les cadenes polipeptídiques, com ara el lligament d'una sonda fluorescent a les cadenes laterals dels aminoàcids.[14] Aquests mètodes són útils en laboratoris bioquímics i de biologia cel·lular, però generalment no per aplicacions comercials. La síntesi química és ineficient per a polipèptids majors de 300 aminoàcids i les proteïnes sintetitzades poden no arribar a assumir la seva forma nativa o estructura terciària. La majoria de mètodes de síntesi procedeixen amb direcció Cterminal a Nterminal de forma contrària a la reacció biològica.
Les propietats de les proteïnes depenen bàsicament de la cadena lateral R dels aminoàcids que la formen. Segons les propietats dels radicals R les propietats de la proteïna seran unes o unes altres. Les propietats principals són la solubilitat, la desnaturalització, l'especificitat i la seva capacitat amortidora.

- 1. Solubilitat: la solubilitat de les proteïnes es deu als radicals R polars dels seus aminoàcids. Com més en conté més soluble és una proteïna. Normalment aquests radicals es dirigeixen cap a l'exterior i són els que formen ponts d'hidrogen amb l'aigua. La polaritat d'aquests radicals es pot veure augmentada en solucions salines diluïdes. Els canvis de PH, que modifiquen el grau de ionització dels radicals R dels aminoàcids, afecten també a la solubilitat de la proteïna.
- 2. Desnaturalització: consisteix en la pèrdua de l'estructura d'una proteïna. Aquesta pèrdua d'estructura es deu al trencament dels enllaços que la constitueixen, principalment causat per un augment de la temperatura, variacions en el PH, canvis de la concentració salina del medi o agitació molecular. Normalment les proteïnes desnaturalitzades adopten una conformació filamentosa i precipiten. Això passa perquè les parts hidrofòbiques de la proteïna poden interaccionar amb altres proteïnes formant grans condensacions. Mentre una proteïna està desnaturalitzada generalment les seves funcions queden anul·lades, tot i que en molts casos, quan es torna a les condicions inicials la proteïna es pot tornar a renaturalitzar.
- 3. Especificitat: moltes proteïnes han d'interaccionar constantment amb altres molècules. Per poder distingir les molècules amb les quals han d'interaccionar de tota la resta presenten una estructura tridimensional i aminoàcids específics. Això els permet dur a terme un control fisiològic molt precís. En són exemples els enzims, les immunoglobulines, la insulina…
- 4. Capacitat amortidora: a causa del caràcter amfòter dels aminoàcids,[16] les proteïnes poden neutralitzar petites variacions del pH d'un medi (solució tampó), cedint o captant protons (H+) segons sigui convenient.
Les proteïnes constituïdes tan sols per aminoàcids s'anomenen holoproteïnes, mentre que si a més d'aminoàcids estan formades per algun tipus de molècula diferent, reben el nom d'heteroproteïnes.
Dins del grup de les holoproteïnes distingim les proteïnes filamentoses i les proteïnes globulars. Les heteroproteïnes o proteïnes conjugades se subdivideixen en cinc grups diferents, segons quin tipus de molècula està associada al seu grup proteic: cromoproteïnes, glicoproteïnes, lipoproteïnes, fosfoproteïnes i nucleoproteïnes.
Holoproteïnes
[modifica]Les proteïnes filamentoses: presenten estructura secundària i majoritàriament tenen funcions estructurals. Són insolubles en aigua i es troben principalment en animals. Hi pertanyen els col·lagens, les α-queratines, les β-queratines o fibroïnes i les elastines.
Les proteïnes globulars: presenten nivells estructurals superiors. Solen formar estructures compactades amb els grups hidròfils orientats cap a l'exterior i els grups hidròfobs dirigits cap a l'interior. Degut a això normalment són dispersables en aigua i solucions salines. Hi pertanyen les protamines, les histones, les prolamines, les glutenines, les albúmines i les globulines (ordenades de menys a més pes molecular).
Heteroproteïnes
[modifica]Les heteroproteïnes són molècules constituïdes per la unió d'un grup proteic amb un altre no proteic mitjançant enllaços forts. El grup no proteic s'anomena grup prostètic. Les heteroproteïnes es classifiquen segons el grup prostètic que les formi.
Cromoproteïnes: són els pigments i tenen com a grup prostètic una substància amb color. Segons el tipus de grup prostètic distingim els pigments porfirínics i els pigments no porfirínics. Els pigments porfirínics tenen com a grup prostètic una porfirina, és a dir, un anell tetrapirròlic, al centre del qual s'hi troba un catió metàl·lic. Si el catió metàl·lic és l'ió ferrós (Fe 2+), la porfirina rep el nom de grup hemo. Són exemples de pigments porfirínics l'hemoglobina, la mioglobina, els citocroms i la catalasa. Els pigments no porfirínics tenen com a grup prostètic una molècula diferent de la porfirina. En són exemples l'hemocianina i l'hemeritrina.
Glicoproteïnes: tenen com a grup prostètic molècules glucídiques. La part glucídica és molt variable, i permet donar especificitat a les molècules. És per això que solen tenir funcions estructurals i de reconeixement. Són exemples de glicoproteïnes les immunoglobulines, les glicoproteïnes de membrana, l'hormona estimulant del fol·licle (FSH) i l'hormona estimulant de la tiroide (TSH), els proteoglicans (quan la part proteica és molt més petita que la glucídica), entre d'altres.
Lipoproteïnes: tenen com a grup prostètic àcids grassos. Solen trobar-se a les membranes citoplasmàtiques, tot i que, a la sang per exemple, realitzen funcions de transport de lípids.
Fosfoproteïnes: tenen com a grup prostètic l'àcid fosfòric (H₃PO₄). Són exemples la caseïna i la vitel·lina.
Nucleoproteïnes: són les heteroproteïnes que tenen un àcid nucleic. Per exemple les associacions de DNA amb histones o protamina per formar la cromatina al nucli cel·lular constitueixen nucleoproteïnes.
Funcions cel·lulars
[modifica]Les proteïnes són els actors principals dins la cèl·lula, executant les missions especificades per la informació codificada als gens.[10] Amb l'excepció de certs tipus d'ARN, la majoria de la resta de molècules biològiques són elements relativament inerts sobre els quals actuen les proteïns. Les proteïnes representen la meitat del pes en sec de la cèl·lula d'escheríchia coli, mentre que altres macromolècules com l'ADN i l'ARN només representen el 3% i el 20%, respectivament.[18] El conjunt de proteïnes expressades en una cèl·lula o un tipus de cèl·lula determinat és el seu proteoma.

La característica principal de les proteïnes, que també permet la seva gran varietat de funcions, és la seva capacitat d'unir-se a altres molècules de manera específica i ferma. La regió de la proteïna que s'encarrega d'unir-se a una altra molècua és coneguda com a lloc d'unió i sol ser una depressió o "butxaca" de la superfície molecular. Aquesta capacitat d'unió és mitjançada per l'estructura terciària de la proteïna, que defineix la butxaca del lloc d'unió, i per les propietats químiques de les cadenes laterals dels aminoàcids adjacents. La unió proteica pot ser extremament ferma i específica; per exemple, la proteïna inhibidor de la ribonucleasa s'uneix a l'angiogenina humana amb una constant de dissociació subfemtomolar (<10−15 M) però no s'uneix gens al seu homòleg dels amfibis onconasa (>1 M). A vegades, canvis químics minúsculs com ara l'addició d'un únic grup metil a una parella d'unió pot ser suficient per eliminar gairebé totalment la unió; per exemple, l'aminoacil ARNt sintetasa específica de l'aminoàcid valina discrimina contra la cadena lateral molt similar de l'aminoàcid isoleucina.[19]
Les proteïnes es poden unir a altres proteïnes a més d'unir-se a substrats de molècules petites. Quan les proteïnes s'uneixen específicament a altres còpies de la mateixa molècula, poden oligomeritzar per formar fibril·les; aquest procés se sol produir en proteïnes estructurals que consisteixen en monòmers globulars que s'autoassocien per formar fibres rígides. Les interaccions proteïna-proteïna també regulen l'activitat enzimàtica, controlen el progrés del cicle cel·lular i permeten l'assemblatge de grans complexos proteics que executen nombroses reaccions relacionades amb una funció biològica comuna. Les proteïnes també poden unir-se, o fins i tot integrar-se, a membranes cel·lulars. La capacitat de les parelles d'unió d'induir canvis conformacionals en les proteïnes permet la construcció de xarxes de senyalització enormement complexes.[20] Cosa important, com que les interaccions entre proteïnes són reversibles, i depenen significativament de la dispnibilitat de grups diferents de proteïnes aparellades per formar agregats que són capaços d'executar conjunts discrets de funcions, l'estudi de les interaccions entre proteïnes específiques és una clau a la comprensió d'aspectes importants del funcionament cel·lular, i en última instància les propietats que distingeixen els tipus concrets de cèl·lules.[21][22]
Nutrició
[modifica]La majoria de microorganismes i plantes poden biosintetitzar els vint aminoàcids estàndard, mentre que els animals (incloent-hi els humans) han d'obtenir-ne alguns de la seva dieta.[18] Els aminoàcids que un organisme no pot sintetitzar per si sol reben el nom d'aminoàcids essencials. Els enzims claus que sintetitzen certs aminoàcids no existeixen en els animals, com per exemple l'aspartocinasa, que catalitza el primer pas de la síntesi de lisina, metionina i treonina a partir de l'aspartat. Si hi ha aminoàcids presents al medi, els microorganismes poden conservar energia assimilant els aminoàcids del seu entorn i subregulant les seves rutes biosintètiques.
Els animals obtenen aminoàcids mitjançant el consum d'aliments que contenen proteïnes. Les proteïnes ingerides són descompostes a través de la digestió, cosa que sol implicar la desnaturalització de les proteïnes per mitjà de l'exposició a àcids i la hidròlisi per part d'enzims anomenats proteases. Alguns aminoàcids ingerits són utilitzats per la biosíntesi de proteïnes, mentre que d'altres són convertits en glucosa a través de la gluconeogènesi, o introduïts al cicle de Krebs. Aquest ús de les proteïnes com a aliment és especialment en condicions d'inanició car permet que el cos utilitzi les seves pròpies proteïnes per mantenir-se en vida, especialment les proteïnes dels músculs.[23] Els aminoàcids també són una important font dietètica de nitrogen.
Història
[modifica]Les proteïnes foren reconegudes com una classe distinta de molècules biològiques al segle xviii per Antoine Fourcroy i altres, que es distingien per la seva habilitat de coagular o flocular sota tractaments amb calor o àcid. Exemples notables d'aquell temps eren l'albúmina de la clara d'ou, l'albúmina de la sang, la fibrina i el gluten del blat. El químic neerlandès Gerhardus Johannes Mulder dugué a terme l'anàlisi elemental de proteïnes comunes i descobrí que gairebé totes les proteïnes tenen la mateixa fórmula empírica: C400H620N100O120P1S1.[24] El terme "proteïna" fou proposat per descriure aquestes molècules el 1838 pel soci de Mulder Jöns Jacob Berzelius; el terme deriva del mot grec πρωτειος (proteios), que significa "al capdavant" o "situat al davant".[25] Més tard, Mulder identificà els productes de la degradació de les proteïnes, com l'aminoàcid leucina, del qual trobà un pes molecular (gairebé correcte) de 131 Da.[24]
La dificultat a l'hora de purificar proteïnes en grans quantitats feu que resultés molt difícil estudiar-les pels primers bioquímics. Per tant, els primers estudis se centraren en proteïnes que es podien purificar en grans quantitats, com ara les de la sang, la clara d'ou, diverses toxines o enzims digestius/metabòlics obtinguts d'escorxadors. En la dècada del 1950 l'Armour & Co. (Hot Dog) purificà un quilogram de ribonucleasa A pancreàtica bovina pura i el posà a lliure disposició dels científics; aquest gest contribuí a fer que la ribonucleasa A esdevingués un objectiu principal de l'estudi bioquímic durant les dècades següents.[24]
Es dona crèdit a Linus Pauling per la predicció reeixida de l'estructura secundària de proteïnes regulars basant-se en els enllaços d'hidrogen, una idea proposada per primera vegada per William Astbury el 1933.[26] El treball posterior de Walter Kauzmann sobre la desnaturalització,[27][28] basat en part en estudis previs de Kaj Linderstrøm-Lang,[29] contribuí a la comprensió del plegament i l'estructura de les proteïnes mediats per interaccions hidrofòbiques. El 1949, Fred Sanger determinà correctament la seqüència d'aminoàcids de la insulina, demostrant així de manera concloent que les proteïnes consistien en polímers lineals d'aminoàcids i no de cadenes ramificades, col·loides o ciclols.[30] Les primeres estructures de resolució atòmica de les proteïnes foren obtingudes per difracció de raigs X a la dècada del 1960 i per NMR a la dècada del 1980. El 2009, el Protein Data Bank compta amb més de 55.000 estructures de resolució atòmica de proteïnes.[31] En temps més recents, la criomicroscòpia electrònica de grans assemblatges macromoleculars[32] i la predicció computada de l'estructura de petits dominis proteics[33] són dos mètodes que s'acosten a la resolució atòmica.
Referències
[modifica]- ↑ Ridley, M. (2006). Genome. Nova York, NY: Harper Perennial. ISBN 0-06-019497-9
- ↑ Maton A., Hopkins J., McLaughlin C. W., Johnson S., Warner M. Q., LaHart D., Wright J. D.. Human Biology and Health. Englewood Cliffs, New Jersey, USA: Prentice Hall, 1993. ISBN 0-13-981176-1. OCLC 32308337.
- ↑ Sumner, J. B. «The isolation and crystallization of the enzyme urease. Preliminary paper». Journal of Biological Chemistry, 69, 1926, pàg. 435–41. Arxivat de l'original el 2007-09-29 [Consulta: 3 maig 2009].
- ↑ Muirhead H, Perutz M. «Structure of hemoglobin. A three-dimensional fourier synthesis of reduced human hemoglobin at 5.5 Å resolution». Nature, 199, 4894, 1963, pàg. 633–38. DOI: 10.1038/199633a0. PMID: 14074546.
- ↑ Kendrew J., Bodo G., Dintzis H., Parrish R., Wyckoff H., Phillips D. «A three-dimensional model of the myoglobin molecule obtained by x-ray analysis». Nature, 181, 4610, 1958, pàg. 662–66. DOI: 10.1038/181662a0. PMID: 13517261.
- ↑ Nelson D. L., Cox M. M. (2005). Lehninger's Principles of Biochemistry, 4th Edition. W. H. Freeman and Company, Nova York.
- ↑ Gutteridge A., Thornton J. M. «Understanding nature's catalytic toolkit». Trends in Biochemical Sciences, 30, 11, 2005, pàg. 622–29. DOI: 10.1016/j.tibs.2005.09.006. PMID: 16214343.
- ↑ Murray et al., p. 19.
- ↑ Murray et al., p. 31.
- ↑ 10,0 10,1 Lodish H., Berk A., Matsudaira P., Kaiser C. A., Krieger M., Scott M. P., Zipurksy S. L., Darnell J. (2004). Molecular Cell Biology 5a edició. WH Freeman and Company: Nova York, NY.
- ↑ Dobson CM. (2000). The nature and significance of protein folding. In Mechanisms of Protein Folding 2nd ed. Ed. RH Pain. Frontiers in Molecular Biology series. Oxford University Press: New York, NY.
- ↑ Fulton A, Isaacs W «Titin, a huge, elastic sarcomeric protein with a probable role in morphogenesis». Bioessays, 13, 4, 1991, pàg. 157-61. PMID: 1859393.
- ↑ Bruckdorfer T., Marder O., Albericio F. «From production of peptides in milligram amounts for research to multi-tons quantities for drugs of the future». Curr Pharm Biotechnol, 5, 1, 2004, pàg. 29-43. PMID: 14965208.
- ↑ Schwarzer D., Cole P. «Protein semisynthesis and expressed protein ligation: chasing a protein's tail». Curr Opin Chem Biol, 9, 6, 2005, pàg. 561-9. PMID: 16226484.
- ↑ ↑ Jimeno A., Ugedo L.. Biologia 1 Batxillerat. Barcelona: Grup Promotor/ Santillana Educación, S.L, 2.008. ISBN 978-84-7918-334-9. Pàg.70
- ↑ ↑ Jimeno A., Ugedo L.. Biologia 1 Batxillerat. Barcelona: Grup Promotor/ Santillana Educación, S.L, 2.008. ISBN 978-84-7918-334-9. Pàg. 72
- ↑ ↑ Jimeno A., Ugedo L.. Biologia 1 Batxillerat. Barcelona: Grup Promotor/ Santillana Educación, S.L, 2.008. ISBN 978-84-7918-334-9. Pàg. 61
- ↑ 18,0 18,1 Voet D., Voet J. G. (2004). Biochemistry Volum 1 3a edició. Wiley: Hoboken, NJ.
- ↑ Sankaranarayanan R., Moras D. «The fidelity of the translation of the genetic code». Acta Biochimica Polonica, 48, 2, 2001, pàg. 323–35. PMID: 11732604.
- ↑ van Holde i Mathews, p. 830–49.
- ↑ Copland J. A., Sheffield-Moore M., Koldzic-Zivanovic N., Gentry S., Lamprou G., Tzortzatou-Stathopoulou F., Zoumpourlis V., Urban R. J., Vlahopoulos S. A.. Sex steroid receptors in skeletal differentiation and epithelial neoplasia: is tissue-specific intervention possible? Bioessays. 2009 Abril del 20. [Epub ahead of print]PMID: 19382224
- ↑ Samarin S, Nusrat A. Regulation of epithelial apical junctional complex by Rho family GTPases. Front Biosci. 2009 Jan 1;14:1129-42. Review. PMID: 19273120
- ↑ Brosnan J. «Interorgan amino acid transport and its regulation». Journal of Nutrition, 133, 6 Suppl 1, 2003, pàg. 2068S–72S. PMID: 12771367.
- ↑ 24,0 24,1 24,2 Perrett D. «From 'protein' to the beginnings of clinical proteomics». Proteomics – Clinical Applications, 1, 8, 2007, pàg. 720–38.
- ↑ Reynolds J. A., Tanford C.. Nature's Robots: A History of Proteins (Oxford Paperbacks). Oxford University Press, USA, 2003, p. 15. ISBN 0-19-860694-X.
- ↑ Pauling L., Corey R. B., Branson H. R. «The structure of proteins: two hydrogen-bonded helical configurations of the polypeptide chain» (PDF). Proceedings of the National Academy of Sciences U.S.A., 37, 1951, pàg. 235–40.
- ↑ Kauzmann W. «Structural factors in protein denaturation». Journal of Cellular Physiology. Supplement, 47, Suppl 1, 1956, pàg. 113–31. PMID: 13332017.
- ↑ Kauzmann W. «Some factors in the interpretation of protein denaturation». Advances in Protein Chemistry, 14, 1959, pàg. 1–63. PMID: 14404936.
- ↑ Kalman S. M., Linderstrom-Lang K., Ottesen M., Richards F. M. «Degradation of ribonuclease by subtilisin». Biochimica et Biophysica Acta, 16, 2, 1955, pàg. 297–99. PMID: 14363272.
- ↑ Sanger F. «The terminal peptides of insulin». Biochemical Journal, 45, 5, 1949, pàg. 563–74. PMC: 1275055. PMID: 15396627.
- ↑ «RCSB Protein Data Bank». Arxivat de l'original el 2015-04-18. [Consulta: 14 abril 2009].
- ↑ Zhou Z. H. «Towards atomic resolution structural determination by single-particle cryo-electron microscopy». Current Opinion in Structural Biology, 18, 2, 2008, pàg. 218–28. DOI: 10.1016/j.sbi.2008.03.004. PMID: 18403197.
- ↑ Keskin O., Tuncbag N., Gursoy A. «Characterization and prediction of protein interfaces to infer protein-protein interaction networks». Current Pharmaceutical Biotechnology, 9, 2, 2008, pàg. 67–76. PMID: 18393863 [Consulta: 14 abril 2009].[Enllaç no actiu]
Vegeu també
[modifica]Bibliografia
[modifica]- Branden C., Tooze J.. Introduction to Protein Structure. Nova York: Garland Pub, 1999. ISBN 0-8153-2305-0.
- Jimeno A., Ugedo L.. Biologia 1 Batxillerat.. Barcelona: Grup Promotor/ Santillana Educación, S.L, 2.008. ISBN 978-84-7918-334-9.
- Murray R. F., Harper H. W., Granner D. K., Mayes P. A., Rodwell V. W.. Harper's Illustrated Biochemistry. Nova York: Lange Medical Books/McGraw-Hill, 2006. ISBN 0-07-146197-3.
- Van Holde K, E., Mathews C. K.. Biochemistry. Menlo Park, Califòrnia: Benjamin/Cummings Pub. Co., Inc, 1996. ISBN 0-8053-3931-0.
Enllaços externs
[modifica]- Joc per entendre el funcionament de les proteïnes (anglès)
- The Protein Databank (anglès)
- Estructura química dels aminoàcids, els pèptids, els polipèptids i les proteïnes (castellà)
| Conceptes generals | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Rutes específiques |
| ||||||||||||||||||