Lèmurs
| Aquest article tracta sobre els animals. Vegeu-ne altres significats a «Lèmur (mitologia)». |
| Lemuriformes | |
|---|---|
 Lèmur de cua anellada (Lemur catta) | |
| Període | |
| Taxonomia | |
| Superregne | Holozoa |
| Regne | Animalia |
| Fílum | Chordata |
| Classe | Mammalia |
| Ordre | Primates |
| Infraordre | Lemuriformes Gray, 1821 |
| Famílies | |
| Distribució | |
 | |
| Endèmic de | |
Els lèmurs (Lemuroidea) són primats estrepsirrins endèmics de l'illa de Madagascar. Deuen el seu nom als lemures, fantasmes o esperits de la mitologia romana, a causa de les estrepitoses vocalitzacions que emeten, els seus ulls brillants i els costums nocturns d'algunes espècies. Tot i que sovint se'ls confon amb primats ancestrals, els primats antropoides (micos, hominoïdeus i humans) no evolucionaren dels lèmurs, tot i que comparteixen trets morfològics i etològics amb els primats basals.
La seva classificació taxonòmica és controvertida i depèn de quin concepte d'espècie es faci servir. Fins i tot es discuteix la taxonomia de nivell més alt. Alguns experts ubiquen la majoria dels lèmurs a l'infraordre Lemuriformes, mentre que altres creuen que aquest clade conté tots els estrepsirrins existents, situant tots els lèmurs a la superfamília Lemuroidea i els lorísids i galàgids a la superfamília Lorisoidea. Arribaren a Madagascar fa aproximadament 62-65 Ma per rafting (en rais de vegetació) en algun moment en què els corrents oceànics n'afavoriren la dispersió cap a l'illa. Des d'aleshores, els lèmurs han evolucionat per enfrontar-se a un medi altament estacional i les seves adaptacions els donen un nivell de diversitat que rivalitza amb el d'altres grups de primats. Fins a l'arribada dels humans a l'illa, fa aproximadament 2.000 anys, existien lèmurs tan grans com un goril·la mascle. Actualment queden unes cent espècies de lèmurs, la majoria descobertes o promogudes al rang d'espècie a partir de la dècada del 1990.
Amb un pes que va des de 30 g fins a 9 kg, els lèmurs comparteixen molts dels trets bàsics dels primats, com mans i peus amb cinc dits i polzes oposables, i ungles en lloc d'urpes (a la majoria d'espècies). Tanmateix, la seva proporció cervell-massa corporal és més petita que en els primats antropoides i, igual que els altres primats estrepsirrins, tenen el «nas humit» (rinari). Els lèmurs són generalment els estrepsirrins més socials i es comuniquen més amb olors i vocalitzacions que amb senyals visuals. Tenen un metabolisme basal relativament baix, molts es reprodueixen estacionalment, tenen períodes de dormància (com hibernació o letargia) i alguns presenten un domini social de les femelles. La majoria s'alimenta d'una àmplia varietat de fruites i fulles, mentre que altres són especialistes. Tot i que moltes tenen dietes similars, espècies diferents comparteixen els mateixos boscos per un procés de diferenciació de nínxol.
La investigació sobre aquests primats se centrà en la seva taxonomia i la recollida d'espècimens durant els segles xviii i xix. Tot i que les observacions aportades pels primers investigadors de camp foren abundants, els estudis moderns sobre l'ecologia i el comportament d'aquests animals no es desenvoluparen fins a les dècades del 1950 i del 1960. Davant les dificultats provocades per la inestabilitat política i la confusió regnant a Madagascar a mitjans de la dècada del 1970, s'aturaren els treballs de camp, però es tornaren a posar en marxa a la dècada del 1980 i suposaren un gran avenç en el coneixement d'aquests primats. Instal·lacions d'investigació com el Duke Lemur Center de la Universitat Duke han ofert una oportunitat d'avançar en el seu estudi en un ambient controlat. Els lèmurs són importants per a la investigació perquè la seva mescla de característiques primitives i trets compartits amb els primats antropoides pot proporcionar un major coneixement sobre l'evolució dels primats i els humans. Tanmateix, moltes espècies de lèmur estan en perill d'extinció a causa de la pèrdua del seu hàbitat i la caça. Tot i que en general les tradicions locals ajuden a protegir els lèmurs i els boscos on viuen, la tala forestal il·legal, la generalització de la pobresa i la inestabilitat política dificulten i soscaven els esforços per la conservació d'aquests primats.
Etimologia
[modifica]Linné, el fundador de la nomenclatura binomial moderna, els donà el nom de lèmurs el 1758, a la 10a edició del seu Systema Naturae. Linné classificà tres espècies sota el gènere Lemur: L. tardigradus (el loris esvelt vermell, actualment Loris tardigradus), L. catta (el lèmur de cua anellada) i L. volans (el lèmur volador de les Filipines, avui en dia Cynocephalus volans).[2] Tot i que el terme «lèmur» s'aplicà inicialment als loris, aviat fou associat a tots els primats endèmics de Madagascar, que han sigut coneguts des d'aleshores com a «lèmurs».[3] El nom prové del llatí lemures,[4][5] espectre o esperits de la mort coneguts com a larvae o lemures a l'antiga Roma.[6] Linné coneixia els hàbits nocturns i l'aspecte fantasmagòric de lèmurs i loris,[7] així com els seus moviments silenciosos de nit, els fantasmagòrics ulls reflectors i les seves crides. També és possible que els pobladors malgaixos li haguessin explicat llegendes que parlen dels lèmurs com les ànimes dels avantpassats.[8] Estant familiaritzat amb les obres de Virgili i Ovidi i les seves referències a la Lemuralia o festival de Lemúria, Linné veié una analogia que encaixava al seu esquema de nomenclatura i donà el nom lemur a aquests primats nocturns.[9]
Història evolutiva
[modifica]Els lèmurs són primats prosimis que pertanyen al subordre Strepsirrhini. Igual que altres primats estrepsirrins, com els loris, potos i gàlags, comparteixen trets basals (plesiomorfies) amb els primers primats, de manera que sovint se'ls confon amb primats ancestrals. Tanmateix, els primats antropoides (micos, hominoïdeus i humans) no evolucionaren dels lèmurs. El lèmur evolucionà de forma independent en el seu aïllament a l'illa de Madagascar.[10] Tradicionalment es considerava que tots els estrepsirrins moderns, incloent-hi els lèmurs, evolucionaren d'uns primats primitius coneguts com a «adapiformes» durant l'Eocè (fa entre 56 i 34 Ma) o el Paleocè (fa entre 65 i 56 Ma),[10][11][12] tanmateix, els adapiformes mancaven d'una disposició especialitzada de les dents, coneguda com a pinta dental, que tenen pràcticament tots els estrepsirrins vivents.[13][14][15] Una hipòtesi més recent suggereix que els lèmurs descendeixen de primats lorisiformes (similars a loris). Aquesta hipòtesi està recolzada per estudis comparatius del gen citocrom b i la presència de la pinta dental dels estrepsirrins en ambdós grups.[15][16] En lloc de ser els avantpassats directes dels lèmurs, els adapiformes podrien haver originat tant els lèmurs com els lorisiformes, una escissió que estaria recolzada per estudis de sistemàtica molecular.[15] Segons els estudis moleculars es creu que la divisió posterior entre lèmurs i loris podria haver tingut lloc fa entre 62 i 65 Ma,[17] tot i que altres proves genètiques i el registre fòssil africà suggereixen estimacions més conservadores de 50 a 55 Ma per aquesta divergència.[18]
Havent estat en algun moment part del supercontinent de Gondwana, l'illa de Madagascar quedà aïllada quan se separà d'Àfrica oriental (~160 Ma), l'Antàrtida (~80-130 Ma) i l'Índia (~80-90 Ma).[19][20] Com que es creu que els primers lèmurs aparegueren a Àfrica fa entre 62 i 65 Ma, haurien d'haver creuat el canal de Moçambic, que té una amplada mínima d'uns 560 km.[15] El 1915, el paleontòleg William Diller Matthew indicà que la biodiversitat de mamífers de Madagascar (incloent-hi els lèmurs) només es podia explicar per esdeveniments esporàdics de rafting (dispersió biològica mitjançant rais de vegetació), en els quals grups molt petits foren transportats des de la propera Àfrica en rais de vegetació enredada que arribaven al mar des dels principals rius;[21] aquesta forma de dispersió biològica podria haver-se produït a l'atzar al llarg de milions d'anys.[15][22] A la dècada del 1940, el paleontòleg George Gaylord Simpson encunyà el terme sweepstakes hypothesis ("hipòtesi de la loteria") per aquests esdeveniments aleatoris.[23] El rafting ha sigut des d'aleshores l'explicació més acceptada de la colonització de Madagascar per part dels lèmurs,[24][25] tot i que fins fa poc es creia que aquest viatge era molt improbable a causa dels forts corrents marins que discorren prop de l'illa.[26] Tanmateix, el gener del 2010, un article demostrà que fa aproximadament 60 Ma tant Madagascar com Àfrica es trobaven 1.650 km al sud de les seves posicions actuals, situant-les en un gir oceànic diferent, i amb corrents que fluïen al contrari de com ho fan en l'actualitat. A més, aquests corrents oceànics eren més forts que avui en dia, de manera que haurien empès els rais de vegetació més ràpidament, escurçant el viatge fins a uns trenta dies o menys, un temps prou curt perquè un petit mamífer pogués sobreviure fàcilment. Quan les plaques continentals derivaren cap al nord, els corrents canviaren gradualment i fa 20 Ma es tancà la finestra oberta per a la dispersió oceànica, aïllant amb eficàcia els lèmurs i la resta de la fauna terrestre malgaixa de la plataforma continental africana.[26] Aïllats a Madagascar amb pocs mamífers competidors, els lèmurs no hagueren de contendre amb altres grups de mamífers arborícoles evolucionats, com ara els esciúrids,[27] ni tampoc tingueren necessitat de competir amb els micos, que evolucionaren més tard i tenien una intel·ligència, una agressivitat i un caràcter evasiu que els conferia un avantatge sobre altres primats en l'explotació del medi.[5][14]
Distribució i diversitat
[modifica]
Els lèmurs s'han adaptat per ocupar molts nínxols ecològics oberts des de la seva arribada a Madagascar.[14][27] El seu nivell de diversitat tant en comportament com en morfologia (aspecte extern) rivalitza amb el d'altres grups de primats existents arreu del món.[5] La seva mida va des dels 30 g del lèmur ratolí de Berthe (Microcebus berthae), el primat més petit del món,[28] fins als 160-200 kg de l'espècie recentment extinta Archaeoindris fontoynonti[29] i han desenvolupat diverses formes de locomoció, diferents formes de complexitat social i adaptacions úniques al clima local.[14][30]
Com que els lèmurs manquen de qualsevol tipus de tret compartit que els distingeixi dels altres primats, la seva diversitat ha ajudat a definir-los.[31] Diferents tipus de lèmurs han desenvolupat combinacions úniques de característiques atípiques per enfrontar-se al clima rigorós i de forts contrasts estacionals de Madagascar, com l'emmagatzematge de greix estacional, l'hipometabolisme (com hibernació o letargia), grups de mida petita, baixa cefalització (mida cerebral relativa), comportament catemeral (amb activitat tant de dia com de nit) i temporades de cria estrictes.[11][30] Es creu que les limitacions extremes per obtenir recursos i la cria estacional han donat lloc a tres altres comportaments relativament comuns en els lèmurs: el domini social de les femelles, el monomorfisme sexual i un model de competició entre mascles per aparellar-se amb baixos nivells d'antagonisme, com ara la competència espermàtica.[32]
Abans de l'arribada dels humans, fa aproximadament 1.500-2.000 anys, els lèmurs es podien trobar arreu de l'illa.[27] Tanmateix, els primers pobladors humans aviat convertiren els boscos en arrossars i prats per mitjà de l'agricultura de tala i crema (coneguda localment com a tavy), restringint els lèmurs a la perifèria de l'illa, aproximadament el 10% de la seva superfície terrestre total, equivalent a uns 60.000 km².[33] Actualment, la diversitat i complexitat de les comunitats de lèmurs augmenta amb la diversitat floral i les precipitacions, particularment a les selves tropicals de la costa oriental, on aquesta diversitat floral i precipitacions són més importants.[12] Malgrat les seves adaptacions per resistir a condicions extremes, la destrucció del seu hàbitat i la caça han fet que les poblacions de lèmurs i la seva diversitat hi hagin disminuït bruscament, causant l'extinció recent d'almenys disset espècies en vuit gèneres,[27][29][34] coneguts col·lectivament com a «lèmurs subfòssils». La majoria d'espècies i subespècies actuals estan amenaçades o en perill d'extinció. Llevat que es produeixi un canvi de tendència, probablement continuaran produint-se extincions.[35]
Fins a temps recents existiren lèmurs de gran mida a Madagascar. Actualment representats només per restes recents o subfòssils, eren formes modernes que en el passat formaren part de la rica diversitat d'aquests primats que evolucionaren en el seu aïllament insular. Algunes de les seves adaptacions eren diferents de les que es poden veure en els seus parents vivents.[27] Disset de les espècies extintes eren més grans que les actuals, algunes amb un pes que arribava fins a 200 kg,[5] i que es creu que podrien haver sigut actives durant el dia.[36] No només diferien dels lèmurs actuals tant en mida com en aspecte, sinó que també ocupaven nínxols ecològics que ja no existeixen o que ara estan deshabitats.[27] Grans extensions de Madagascar que ara manquen de boscos i de lèmurs, en un temps albergaren comunitats diverses de primats que incloïen més de vint espècies de lèmur que abastaven l'ample rang de mida d'aquests animals.[37]
Classificació taxonòmica i filogènesi
[modifica]| Filogènesi dels lèmurs;[24][38][37] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Des d'un punt de vista taxonòmic, el gènere (biologia) Lemur, que s'associà originalment al terme «lèmur», actualment només conté el lèmur de cua anellada (Lemur catta). Actualment, «lèmur» és el nom comú que s'utilitza per referir-se a tots els primats malgaixos.[39]
La taxonomia dels lèmurs és controvertida i no tots els experts estan d'acord, en bona part a causa del recent augment en el nombre d'espècies reconegudes.[31][40][41] Segons Russell Mittermeier, president de Conservation International (CI), el taxonomista Colin Groves i altres científics, hi ha unes cent espècies o subespècies de lèmurs reconegudes que perviuen en l'actualitat, dividides en cinc famílies i quinze gèneres.[42] Com que les dades genètiques indiquen que els lèmurs subfòssils (extints recentment) estaven estretament relacionats amb els actuals,[43] es poden incloure unes tres famílies, vuit gèneres i disset espècies addicionals en el total.[29][34] Tanmateix, altres experts qualifiquen aquestes afirmacions d'«inflació taxonòmica»[41] i redueixen el nombre d'espècies vivents a cinquanta.[31]
També hi ha discrepàncies sobre la classificació dels lèmurs dins el subordre Strepsirrhini, tot i que la majoria d'experts coincideixen en el mateix arbre filogenètic. En una taxonomia publicada per Colin Groves, l'ai-ai fou situat dins el seu propi infraordre, Chiromyiformes, mentre que la resta de lèmurs foren classificats a Lemuriformes.[44] En una altra taxonomia, Lemuriformes inclou tots els estrepsirrins vivents en dues superfamílies, Lemuroidea per tots els lèmurs i Lorisoidea pels loris i gàlags.[18]
| 3 infraordres, 2 superfamílies[44] | 1 infraordre, 2 superfamílies[18][45] |
|---|---|
|
|

La taxonomia dels lèmurs ha canviat considerablement des de la primera classificació taxonòmica realitzada per Carl von Linné el 1758. Un dels reptes més grans fou la classificació de l'ai-ai, que ha sigut objecte de debat fins molt recentment.[5] Fins que Richard Owen publicà un estudi anatòmic definitiu el 1866, els primers naturalistes no estaven segurs si l'ai-ai (gènere Daubentonia) era un primat, un rosegador o un marsupial[46][47][48] i la seva ubicació entre els primats també fou problemàtica fins fa molt poc temps. Basant-se en la seva anatomia, els investigadors han trobat motius per classificar el gènere Daubentonia com un índrid especialitzat, com a grup germà dels estrepsirrins i com a tàxon de posició indefinida dins l'ordre dels primats.[16] Les proves moleculars han demostrat que Daubentoniidae és basal respecte a tots els lemuriformes[16][49] i, el 2008, Russell Mittermeier, Colin Groves i altres no tingueren en compte la seva localització en una taxonomia de nivell superior i definiren els lèmurs com a grup monofilètic amb cinc famílies vivents, incloent-hi Daubentoniidae.[42]
Les relacions entre famílies també han estat problemàtiques i encara estan pendents de ser resoltes definitivament.[16] Per complicar encara més la qüestió, diversos fòssils de primats del Paleogen de fora de Madagascar, com ara Bugtilemur, han sigut classificats com a lèmurs.[50] Tanmateix, el consens científic no accepta aquestes classificacions basades en proves genètiques,[16][49] per la qual cosa generalment es considera que els primats malgaixos són monofilètics.[16][24][38] Una altra àrea controvertida és la relació entre els lepilemúrids i el tàxon extint dels megaladàpids, anteriorment agrupats an la mateixa família a causa de les semblances en la seva dentadura,[51] però si es tenen en compte els estudis genètics, ja no es considera que estiguin estretament relacionats.[38][52]
S'han fet molts canvis taxonòmics a escala de gènere, tot i que aquestes revisions han resultat més concloents, sovint sostingudes per anàlisis genètiques i moleculars. Una de les revisions més notables consistí en la divisió gradual d'un ample gènere Lemur en gèneres separats pel lèmur de cua anellada, els lèmurs de collar i els lèmurs bruns (també anomenats lèmurs autèntics), a causa d'un gran nombre de diferències morfològiques.[53][54]
Després de les revisions taxonòmiques de Mittermeier, Groves i altres, el nombre d'espècies reconegudes ha passat de trenta-tres espècies i subespècies el 1994 a aproximadament cent el 2008.[31][42][55] Si es continua amb les investigacions citogenètiques i de genètica molecular, a més dels estudis de camp en curs, en particular amb espècies críptiques com ara els lèmurs ratolí, el nombre d'espècies reconegudes probablement seguirà creixent.[31] Tanmateix, l'augment en el nombre d'espècies reconegudes ha tingut també els seus crítics entre els estudiosos dels lèmurs i els taxonomistes. També cal considerar que, al cap i a la fi, les classificacions depenen del concepte d'espècie que es faci servir i alguns especialistes proposen que es realitzin anàlisis més minucioses.[31][41]
Anatomia i fisiologia
[modifica]Els lèmurs són un grup divers en termes morfològics i fisiològics.[31] Alguns, com ara els lepilemúrids i els índrids, tenen les extremitats inferiors més llargues que les superiors i són excel·lents saltadors.[56][57][58] Com a folívors que són, els índrids compten amb un aparell digestiu especialitzat, amb glàndules salivals molt desenvolupades, un estómac espaiós i un cec allargat que facilita la fermentació.[12][14][59] [60][61] El lèmur nan d'orelles peludes té una llengua molt llarga que li permet alimentar-se de nèctar,[47] i el lèmur de panxa vermella té una llengua similar a un raspall, també especialment adaptada per alimentar-se de nèctar i pol·len.[12] L'ai-ai ha desenvolupat algunes característiques que són úniques entre els primats i que el distingeixen dels altres lèmurs. Aquests trets diferenciadors inclouen unes incisives de creixement continu, igual que les dels rosegadors, que li permeten rosegar fusta i llavors de closca dura, un dit mitjà filiforme (molt fi, en forma de filament) per extreure menjar de forats diminuts, unes grans orelles similars a les dels ratpenats per detectar buits als arbres,[14][27][47][62] o l'ús de senyals acústics per buscar aliment.[46]
Compten amb una enorme varietat de mides, incloent-hi el primat més petit del món i, fins fa poc, alguns dels més grans. Actualment, la seva mida va des dels 30 g del lèmur ratolí de Berthe fins als 7-9 kg de l'indri i el sifaca de diadema,[63][59] però si es tenen en compte les espècies recentment extintes, la seva gamma de mides s'estén fins a l'equivalent al d'un goril·la mascle, amb els 160-200 kg d'Archaeoindris fontoynonti.[5][29]

Com tots els primats, els lèmurs tenen cinc dits divergents dotats d'ungles (en la majoria de casos) a les mans i els peus. La majoria tenen una ungla allargada i contreta lateralment, anomenada urpa d'empolainament al segon dit del peu, la qual utilitzen per rascar-se i empolainar-se.[47][64] A més d'aquesta urpa d'empolainament, els lèmurs presenten altres trets característics dels primats estrepsirrins, com ara el rinari (o «nas humit», principal diferència amb els haplorrins), un òrgan vomeronasal totalment funcional que detecta feromones, una barra òssia postorbital i la cavitat ocular sense tancament postorbital, símfisi mandibular no fusionada (mandíbules esquerra i dreta separades), i una proporció cervell-massa corporal baixa.[15][65][66]
Els trets compartits amb altres primats prosimis també inclouen un úter bicorne (amb dues banyes, en forma de Y) i la placentació epitelicorial.[13][65] Com que els seus polzes només són pseudooposables, cosa que fa que es moguin de manera menys independent dels altres dits,[64] són menys hàbils a l'hora d'agafar i manipular objectes.[20] L'hàl·lux (primer dit del peu) està molt desenvolupat, cosa que fa més fàcil agafar-se a les branques dels arbres.[47] Un error bastant comú és la creença que els lèmurs tenen una cua prènsil; tanmateix, és una característica que entre els primats només es dona en els micos del Nou Món i, més concretament, els atèlids.[64] Els lèmurs compten amb un sentit de l'olfacte molt desenvolupat, un tret compartit amb la majoria dels altres mamífers i primats primitius, però no amb els primats superiors, que s'orienten principalment amb la vista.[20]
Els lèmurs són atípics per la gran varietat de les estructures socials que tenen, tot i que generalment manquen de dimorfisme sexual quant a la mida i la morfologia de les dents canines.[12][39] Tanmateix, en algunes espècies les femelles solen ser més grans,[46] i dues espècies del gènere Eulemur), el lèmur de cap gris i el lèmur bru de front vermell, presenten diferències de mida a les canines.[67] Entre els lèmurs bruns es dona un cert dicromatisme sexual (diferències entre els sexes en la coloració de la pell),[39] tot i que aquesta diferència varia des de sorprenentment òbvia, com en el lèmur negre, a gairebé imperceptible en el lèmur bru.[67]
S'ha descobert recentment que hi ha lèmurs que formen complexos críptics d'espècies, especialment entre dels lèmurs mostela i lèmurs ratolí. Les subespècies de lèmurs mostela se solien definir basant-se en petites diferències morfològiques, tot i que noves proves genètiques han recolzat l'estatus d'espècie pròpia atorgat a diverses poblacions regionals.[52] En el cas dels lèmurs ratolí, el lèmur ratolí gris, el lèmur ratolí de Ravelobe i el lèmur ratolí de Goodman fa poc eren considerats com una única espècie, fins que foren identificats com a espècies críptiques per proves genètiques.[68]
Dentadura
[modifica]| Família | Fórmula dental caduca [51][69] |
Fórmula dental permanent [39][47][70][71] |
|---|---|---|
| Cheirogaleidae, Lemuridae | | align=center| | |
| Lepilemuridae | | align=center| | |
| Archaeolemuridae † | | align=center| | |
| Megaladapidae † | | align=center| | |
| Indriidae, Palaeopropithecidae † | [N 1] | [N 2] |
| Daubentoniidae | | align=center| |
Els lèmurs són animals heterodonts (tenen diferents tipus de dents) i la seva dentadura deriva d'una dentadura permanent ancestral de . Els índrids, lèmurs mostela, l'ai-ai i els extints Mesopropithecus, Archaeolemuridae i Megaladapis han reduït la seva dentadura i han perdut incisives, canines o premolars.[73] La dentadura caduca (dents de llet) ancestral és , però els índrids joves, els ai-ais, Megaladapis, els paleopropitècids i, probablement, els arqueolemúrids tenen menys dents caduques.[51][69]
També hi ha diferències significatives en la morfologia i topografia dental dels lèmurs. L'indri per exemple, té dents que estan perfectament adaptades per arrancar fulles i esclafar llavors.[59] A la pinta dental de la majoria de lèmurs, les incisives i canines inferiors són procumbents (cap endavant) i estan alineades i espaiades amb precisió, de manera que serveixen com a eina per l'empolainament i l'alimentació.[15][51][73] Per exemple, els indris no utilitzen la pinta dental només per empolainar-se, sinó també per extreure les grans llavors del resistent epicarpi dels fruits de Beilschmiedia,[74] mentre que els lèmurs forcats utilitzen la seva pinta dental relativament llarga per esquinçar l'escorça dels arbres i aconseguir que en flueixi la saba.[47] Només l'ai-ai, l'extint ai-ai gegant i el més gran dels també extints mesopropitecs manquen de pinta dental funcional, característica dels estrepsirrins.[73][71] En el cas de l'ai-ai, la morfologia de les seves incisives caduques, que cauen poc després del naixement, indica que els seus avantpassats tenien una pinta dental. Perden aquestes dents de llet poc després de néixer[75] i les canvien per unes incisives que creixen contínuament durant tota la vida.[73]
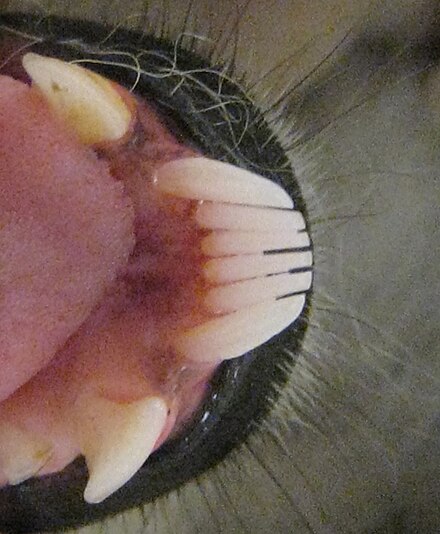
La pinta dental dels lèmurs està formada generalment per sis dents (quatre incisives i dues canines), tot i que en el cas dels índrids, els lèmurs mico i alguns lèmurs peresós l'estructura està formada per quatre dents.[15][73] Com que l'ullal inferior forma part de la pinta dental o s'ha perdut, la dentadura inferior pot ser difícil d'interpretar, sobretot des de la primera premolar (P2) que sovint és caniniforme (té forma de canina) per ocupar el paper de la canina.[51] En els lèmurs folívors, tret dels índrids, les incisives superiors estan absents o són molt reduïdes.[51][73] Utilitzat juntament amb la pinta dental del maxil·lar inferior, aquest complex dental recorda al coixinet dental dels ungulats.[73]
Els lèmurs són atípics entre els primats pel seu ràpid desenvolupament dental, en particular entre les espècies més grosses. Els índrids tenen un creixement corporal relativament lent, però la formació i erupció dentals són molt ràpides.[76] En canvi, els primats simiformes experimenten un desenvolupament dental més lent i tenen amb mida més gran i un desenvolupament morfològic també més lent.[73] Els lèmurs també són dentalment precoços en néixer i ja tenen la dentadura permanent completa al moment del deslletament.[30]
Generalment tenen un esmalt dental fi comparat amb el dels primats antropoides. Això pot causar més desgast i el trencament de les dents anteriors a causa del seu ús intensiu en l'empolainament, l'alimentació i la lluita. No hi ha gaire informació disponible sobre la salut dental dels lèmurs, a part del fet que els lèmurs de cua anellada salvatge de la reserva privada de Berenty (Madagascar) a vegades presenten abscessos a les canines superiors i càries, possiblement a causa del consum d'aliments no nadius.[73]
Sentits
[modifica]
El sentit de l'olfacte és molt important per aquests primats i sovint l'utilitzen per comunicar-se.[12][14][20] Els lèmurs tenen el musell llarg (a diferència del curt musell dels haplorrins), que tradicionalment ha estat vist com una adaptació evolutiva per aconseguir una millor posició del nas per sentir les olors,[14] tot i que un musell llarg no necessàriament es tradueix en una alta agudesa olfactiva, car no és la mida relativa de la fossa nasal el que es relaciona amb l'olor, sinó la densitat de receptores olfactius.[77][78] En canvi, els musells llargs poden facilitar una millor masticació.[78]
El rinari (o «nas humit») és un tret compartit amb altres estrepsirrins i molts altres mamífers, però no amb els primats haplorrins.[47] Tot i que a vegades es diu que potencia el sentit de l'olfacte,[65] en realitat és un òrgan sensorial basat en el tacte que s'uneix amb un òrgan vomeronasal ben desenvolupat. Com que les feromones generalment són molècules grans i no volàtils, el rinari s'utilitza per tocar un objecte marcat per l'olor i transferir les molècules de les feromones baixant pel filtre fins a l'òrgan vomeronasal via els conductes nasopalatins que discorren a través del forat incisiu del paladar dur.[13]
Per comunicar-se mitjançant l'olor, cosa que resulta molt útil de nit, els lèmurs marquen el seu territori amb orina i les glàndules odoríferes situades als canells, la cara interna del colze, les zones genitals i el coll.[13][65] La pell de l'escrot de la majoria de lèmurs mascles té glàndules odoríferes.[79] Els lèmurs de collar i els sifaques mascles tenen una glàndula a la base del coll,[13][47] mentre que el lèmur de nas ample i el lèmur de cua anellada tenen glàndules a les potes anteriors, prop de l'axil·la.[13] Els mascles dels lèmurs de cua anellada també tenen glàndules odoríferes a la cara interna dels avantbraços, juntament amb un esperó similar a una espina, que utilitzen per excavar i alhora impregnar branques d'arbres amb senyals odorífers.[47] També es freguen la cua entre els avantbraços i després prenen part en «lluites pudents», agitant la cua davant dels seus oponents.[13]
| Lèmurs | Primats antropoïdeus | |
|---|---|---|
| Angle entre els ulls | 10-15° | 0° |
| Camp binocular | 114-130° | 140-160° |
| Camp combinat (binocular + perifèric) |
250-280° | 180-190° |
Es considera que els lèmurs (i els estrepsirrins en general) s'orienten visualment en menor mesura que els primats «superiors», car es basen fonamentalment en el seu sentit de l'olfacte i la percepció de feromones. Tenen poc desenvolupada la fòvea de la retina, que confereix una major agudesa visual. Es creu que el septe postorbital (tancament ossi darrere de l'ull) dels primats haplorrins estabilitza lleugerament l'ull, cosa que permet l'evolució de la fòvea; només amb la barra postorbital, els lèmurs han sigut incapaços de desenvolupar la fòvea.[81] Per això, independentment del seu model d'activitat (nocturn, catemeral o diürn), els lèmurs tenen una baixa agudesa visual i una elevada sumació retinal.[30] Tanmateix, els lèmurs tenen un camp visual més ample que els primats antropoides, gràcies a una petita diferència en l'angle entre els ulls, com es pot veure a la taula de l'esquerra.[80]
Tot i que manquen de fòvea, alguns lèmurs diürns tenen una àrea central (porció de la retina que envolta la fòvea i on es produeix la major fotorecepció) rica en cons, tot i que menys agrupada.[81] La proporció bastons-cons d'aquesta àrea central és alta en moltes de les espècies diürnes estudiades, mentre que els antropoïdeus diürns no tenen bastons a la fòvea. Això també suggereix que l'agudesa visual dels lèmurs és inferior a la dels primats antropoides.[82] A més, la proporció bastons/cons varia fins i tot entre espècies diürnes. Per exemple, el sifaca de Verreaux i l'indri tenen pocs cons grans dispersos al llarg d'una retina on predominen els bastons. Els ulls del lèmur de cua anellada tenen un con per cada cinc bastons. D'altra banda, els lèmurs nocturns, com els lèmurs ratolí i els lèmurs nans, tenen les retines compostes únicament de bastons.[13]
Com que són els cons els que fan possible la percepció del color, el predomini dels bastons en els ulls dels lèmurs suggereix que no han desenvolupat la visió en color.[13] S'ha comprovat que el lèmur més estudiat, el de cua anellada, té la visió blava-groga, però no és capaç de distingir matisos vermells i verds.[83] A causa del polimorfisme de les molècules d'opsines, que codifiquen la receptivitat en color, la visió tricromàtica es pot donar molt rarament en les femelles d'algunes espècies, com el sifaca de Coquerel i el lèmur de collar vermell. La majoria de lèmurs són, per tant, monocromàtics o dicromàtics.[13]

La majoria de lèmurs tenen un tapetum lucidum, una capa reflectora de teixits situada als ulls de molts animals vertebrats,[39] però no els primats haplorrins. La seva presència augmenta la capacitat visual dels lèmurs.[30][82] El tapetum coroïdal dels estrepsirrins és únic entre els mamífers perquè es compon de riboflavina cristal·lina. La dispersió òptica resultant és el que en limita l'agudesa visual.[82] Tot i que es creu que tots els lèmurs tenen tapetum, pot haver-hi excepcions al gènere Eulemur, com ara el lèmur negre i el lèmur bru, així com al gènere Varecia.[13][30][82] Tanmateix, com que la riboflavina del tapetum tendeix a dissoldre's i desaparèixer quan és tractada per sotmetre-la a un examen histològic, aquestes excepcions encara són qüestionables.[13]
Metabolisme
[modifica]Els lèmurs tenen un metabolisme basal baix que els ajuda a conservar energia durant l'estació seca, quan escassegen l'aigua i el menjar.[12][58] Poden optimitzar l'ús d'energia baixant el ritme metabòlic a un 20% per sota dels valors corresponents a mamífers amb una massa corporal similar.[84] Per exemple, s'ha observat que el lèmur mostela de cua vermella té un dels metabolismes basals més baixos de tots els mamífers. El seu baix ritme metabòlic pot estar relacionat amb la seva dieta generalment folívora i la seva massa corporal relativament petita.[58] Els lèmurs presenten adaptacions de comportament per complementar aquesta característica, com ara prendre el sol, seure encongits, arraulir-se en grups i compartir llit, per a reduir la pèrdua de calor i conservar energia.[84] Els lèmurs nans i els lèmurs ratolí tenen cicles estacionals de dormància per conservar energia.[84] Abans de l'estació seca acumulen greix al teixit adipós blanc localitzat a la base de la cua i les potes posteriors, fins a doblar el seu pes.[28][85][86] Al final de la temporada seca, la seva massa corporal pot baixar fins a la meitat del que era abans de la temporada.[28] Els que no experimenten estats de dormància també són capaços de suspendre algunes funcions metabòliques. per conservar energia.[84]
Comportament
[modifica]El comportament dels lèmurs és tan variable com la seva morfologia. Les diferències de dieta, sistema social, model d'activitat, locomoció, comunicació, tàctiques per evitar els depredadors, sistema reproductor i nivell d'intel·ligència ajuden a definir els tàxons de lèmurs i distingir espècies. Tot i que freqüentment es diferencien els lèmurs més petits i nocturns dels que són més grans i diürns, sovint hi ha excepcions que ajuden a exemplificar la naturalesa única i diversa d'aquests primats malgaixos.
Dieta
[modifica]
Les seves dietes són molt variades i demostren un alt grau de versatilitat,[87] tot i que la tendència general és que les espècies més petites consumeixin principalment fruita i insectes (és a dir són omnívores), mentre que les espècies més grosses són més herbívores i consumeixen sobretot matèria vegetal.[36] Com tots els primats, els lèmurs afamats mengen gairebé qualsevol cosa que sigui comestible, encara que no sigui un dels seus aliments preferits.[13] Per exemple, el lèmur de cua anellada menja insectes i petits vertebrats quan és necessari,[36][53] per la qual cosa habitualment se'l considera un omnívor oportunista.[73] El lèmur nan de Coquerel és principalment frugívor, però consumeix secrecions dels insectes durant l'estació seca.[36]
Una suposició habitual en mastologia és que els petits mamífers no poden subsistir amb una alimentació exclusivament vegetal i necessiten una dieta rica en calories per sobreviure. Per això es considerava que la dieta dels primats menuts havia de tenir un alt contingut d'insectes (és a dir, serien insectívors), que són rics en proteïnes. Les investigacions han mostrat, tanmateix, que els lèmurs ratolí, els primats vivents més petits, consumeixen més fruita que insectes, cosa que contradiu la hipòtesi general.[13][36]
La matèria vegetal és el component fonamental de la dieta de la majoria d'espècies de lèmur. Espècies d'almenys 109 de totes les famílies de plantes conegudes a Madagascar (un 55%) són aprofitades pels lèmurs. Com que són fonamentalment arborícoles, la majoria d'aquestes espècies vegetals aprofitades són plantes llenyoses, com ara arbres, arbustos o lianes. El lèmur de cua anellada, els lèmurs del bambú i el lèmur de collar són els únics lèmurs que se sap que consumeixen herbes. Tot i que Madagascar és ric en una gran varietat de falgueres, els lèmurs gairebé mai no se les mengen; una possible raó podria ser el fet que les falgueres manquen de les flors, fruites o llavors comuns en la seva dieta. A més, creixen a terra, mentre que els lèmurs passen gran part del temps als arbres. D'altra banda, les falgueres tenen un gust desagradable a causa de l'alt contingut de tanins de les frondes. Igualment, els lèmurs rarament aprofiten els mangles, també a causa del seu alt contingut en tanins.[87] Tanmateix, sembla que alguns lèmurs han desenvolupat respostes contra defenses comunes de les plantes, com ara tanins i alcaloides.[74] El lèmur daurat, per exemple, s'alimenta del bambú gegant Cathariostachys madagascariensis, que conté nivells alts de cianur. Aquest lèmur pot consumir cada dia dotze vegades la dosi habitualment letal per a la majoria de mamífers. Es desconeixen els mecanismes fisiològics que el protegeixen.[12] Al Duke Lemur Center de la Universitat Duke (Estats Units), els lèmurs que vaguen pels recintes a l'aire lliure han sigut observats menjant heura metzinosa (Taxicodendron radicans) sense presentar símptomes d'enverinament.[64]
Molts lèmurs que mengen fulles solen fer-ho durant èpoques d'escassetat de fruita i a vegades perden pes per aquest motiu.[88] La majoria d'espècies de lèmurs, incloent-hi la majoria dels més petits i excloent-ne alguns indris, s'alimenta predominantment de fruita (són frugívores) si n'hi ha. En total, s'ha comprovat que els lèmurs consumeixen la fruita d'almenys 86 famílies de plantes natives i 15 famílies de plantes foranes. Igual que en la majoria de frugívors tropicals, en la seva dieta predomina la fruita de les espècies de Ficus (figueres).[87] En molts primats antropoides, la fruita és una font primària de vitamina C, però a diferència d'ells, els lèmurs (i tots els estrepsirrins) poden sintetitzar la seva pròpia vitamina C.[89] Històricament s'ha considerat que les dietes dels lèmurs en captivitat riques en fruites amb molta vitamina C causaven hemosiderosi, un trastorn provocat per un excés sistèmic de ferro, car la vitamina C incrementa l'absorció d'aquest metall. Tot i que s'ha comprovat que els lèmurs en captivitat són susceptibles a l'hemosiderosi, la freqüència de la malaltia varia d'una institució a l'altra i pot dependre de la dieta, els protocols de manteniment i el patrimoni gènic. Les suposicions sobre aquest problema s'han de posar a prova per cada espècie diferent.[90] Sembla que el lèmur de cua anellada, per exemple, és menys susceptible a aquesta malaltia que altres espècies de lèmur.[91]
Es creu que només vuit espècies de lèmur s'alimenten de llavors (són granívores), tot i que aquesta xifra podria ser inferior a la real, car la majoria d'observacions només mostren el consum de fruita i no indiquen si també es mengen les llavors. Entre aquests lèmurs granívors hi ha diversos índrids, com ara el sifaca de diadema, el sifaca de Tattersall, l'indri[12][61] i l'ai-ai. L'ai-ai, que s'especialitza en espècies amb defenses estructurals, pot mastegar llavors de Canarium, que són més dures que les que són capaços d'obrir els micos del Nou Món.[46] Almenys 36 gèneres de 23 famílies vegetals formen part de la dieta dels lèmurs granívors.[87]
Les inflorescències d'almenys 60 famílies de plantes són consumides per lèmurs amb mides que van des dels diminuts lèmurs ratolí fins als lèmurs de collar, que són relativament grans. Si no utilitzen les flors, a vegades consumeixen el nèctar (són nectarívors) juntament amb el pol·len (pol·linívors). Almenys 24 espècies nadiues de 17 famílies de plantes formen part de la dieta dels lèmurs pel seu pol·len o nèctar.[87]
L'escorça i els exsudats de les plantes, com ara la saba d'arbre, són consumides per poques espècies. La ingestió d'exsudats ha sigut observada amb 18 espècies de plantes i només a les regions seques del sud i l'oest de Madagascar. El lèmur forcat de Masoala i el lèmur nan de Coquerel són les úniques espècies que consumeixen saba d'arbres amb regularitat. Mai no s'ha observat que l'escorça sigui un element significatiu en la dieta dels lèmurs, però almenys quatre espècies en mengen: l'ai-ai, el lèmur mostela de cua vermella, el lèmur bru i el sifaca de Verreaux. L'alimentació amb escorça generalment acompanya l'alimentació amb exsudats, tret del consum per part de l'ai-ai d'escorça d'Afzelia bijuga a la reserva de Nosy Mangabe, al nord-est de l'illa.[87]
També s'ha observat el consum de terra (geofàgia), probablement per facilitar la digestió, obtenir minerals i sals i neutralitzar les toxines. S'han observat sifaques menjant terra de monticles de tèrmits, possiblement per aconseguir una flora intestinal beneficiosa que els ajudi a digerir la cel·lulosa de la seva dieta folívora.[64]
Els lèmurs són animals socials i viuen en grups que en general tenen almenys quinze individus.[12] Entre els seus models d'organització social, es poden mencionar el model «solitari, però social» i el model de «fissió-fusió»,[N 3] «unions de parella» i «grups multimascle».[93] Els lèmurs nocturns són fonamentalment solitaris però socials. De nit busquen aliment totss sols, però en general es reuneixen en grups durant el dia. El grau de socialització varia en funció de l'espècie, el gènere, la localització i l'estació.[27][36] Per exemple, en moltes espècies nocturnes les femelles, juntament amb les seves cries, comparteixen llocs de cria amb altres femelles i possiblement un mascle. Aquests llocs de cria abasten un ample territori i els poden compartir diversos grups de femelles. En el cas dels lèmurs mostela i els lèmurs forcats, una o dues femelles poden compartir un territori, possiblement amb un mascle. A més de compartir espais vitals, també es relacionen vocalment o física amb el seu company de territori mentre busquen aliment durant la nit.[36] Entre els lèmurs diürns es donen molts dels sistemes socials vistos en micos i hominoïdeus.[12][36] Viuen en grups socials relativament permanents i cohesionats. Els grups multimascle són el sistema més comú entre ells, igual que en la majoria de primats antropoïdeus.[N 4] Els lèmurs bruns o vertaders utilitzen aquest sistema social i sovint viuen en grups de deu individus o menys. S'ha comprovat que els lèmurs de collar viuen en societats de fissió-fusió[36] i que els indris formen unions de parella.[93]

Sistemes socials
[modifica]En alguns lèmurs s'ha observat filopatria (la tendència a romandre al seu territori natal o a tornar-hi després d'haver-lo abandonat) entre les femelles, que romanen al seu territori, mentre que els mascles emigren en arribar a la maduresa. En altres espècies, ambdós sexes emigren.[12] En alguns casos, la filopatria femenina pot ajudar a explicar l'evolució de grups multimascle de femelles relacionades, com en el cas del lèmur de cua anellada, el sifaca de Milne-Edwards i el sifaca de Verreaux. Els seus avantpassats podrien haver sigut més solitaris, amb femelles que vivien en parelles mare-fill (díada). Amb el temps, aquestes díades es podrien haver unit amb altres parelles mare-fill veïnes per a defensar recursos dispersos en un territori ample. Si fos així, els grups multimascle dels lèmurs estarien clarament diferenciats en la seva estructura interna en comparació amb els catarrins.[95]
La presència d'un domini social de les femelles diferencia els lèmurs de la majoria dels altres primats i, fins i tot, dels altres mamífers.[12][36][39][96] En la majoria de societats de primats, els mascles són els dominants llevat que les femelles s'uneixin per formar coalicions que els desplacin.[97] Tanmateix, hi ha espècies que no presenten aquest domini femení, com és el cas de moltes espècies d'Eulemur,[36][67] o el lèmur de nas ample.[98] Quan les femelles són les dominants dins d'un grup, la manera de la qual mantenen el domini varia. Els mascles de lèmur de cua anellada actuen de manera submisa amb senyals d'agressió per part de les femelles o sense. En canvi, els mascles de lèmur coronat només actuen de manera submisa quan les femelles es comporten de forma agressiva envers ells. L'agressió de les femelles sovint, però no sempre, està relacionada amb l'alimentació.[99]
Moltes hipòtesis han intentat explicar per què els lèmurs han desenvolupat un domini social de les femelles, a diferència d'altres primats amb estructures socials similars,[12][96] però no s'ha assolit cap consens després de dècades d'investigació. El punt de vista dominant a la bibliografia científica és que el domini femení és una característica avantatjosa a causa dels alts costos de la reproducció i l'escassetat de recursos disponibles,[96] car s'ha comprovat que el domini de les femelles està lligat a un increment de la cura i l'atenció maternals.[97] Tanmateix, quan els costos reproductius i una extrema estacionalitat dels recursos foren comparats amb la mateixa situació entre altres primats, es comprovà l'existència d'un domini masculí en condicions que eren similars o fins i tot més difícils que les condicions a les quals s'enfrontaven els lèmurs. El 2008 una nova hipòtesi revisà aquest model utilitzant la teoria de jocs. Se sostingué que quan dos individus tenien una capacitat de lluita similar, el més necessitat guanyaria el conflicte perquè hi tindria més a perdre. Per això, la femella, amb més necessitats de recursos a causa de l'embaràs, la lactància i la cura materna, tindria més probabilitats de vèncer en conflictes per obtenir recursos amb mascles de la mateixa mida. Tanmateix, aquesta hipòtesi pressuposava l'existència d'un monomorfisme entre sexes.[96] L'any següent es proposà una nova hipòtesi per explicar el monomorfisme, que afirmava que, com que la majoria de femelles de lèmurs només estan sexualment receptives durant un o dos dies cada any, els mascles poden utilitzar una forma més passiva de protecció de la parella: el tap de copulació (una secreció gelatinosa utilitzada en l'aparellament d'alguns animals), que bloqueja el tracte reproductiu de la femella i impedeix que altres mascles s'hi aparellin, reduint la necessitat de comportaments agressius i alentint el camí evolutiu cap al dimorfisme sexual.[32]

En general, els nivells agonístics (o agressius) tendeixen a estar relacionats amb la longitud relativa de les canines. El lèmur de cua anellada té les dents canines superiors llargues i afilades en ambdós sexes i mostra alts nivells d'agonisme. En canvi, l'indri té ullals més petits i mostra nivells inferiors d'agressió.[30] Quan grups veïns de la mateixa espècie defensen els seus respectius territoris, el conflicte pot prendre la forma de defensa ritualitzada. En els sifaques, aquests combats ritualitzats impliquen mirar-se fixament els uns als altres, grunyir, deixar marques d'olor i saltar per ocupar certes seccions de l'arbre, mentre que l'indri defensa el seu territori amb batalles rituals de «cant».[12]
Com altres primats, els lèmurs utilitzen l'empolainament social per alleujar relacions tenses i consolidar relacions. S'empolainen en saludar-se, quan es desperten, en anar a dormir, entre mares i fills, en les relacions juvenils i per fomentar relacions sexuals.[100] A diferència dels primats antropoïdeus, que aparten el pelatge amb les mans i agafen amb els dits o la boca les partícules, com ara pell morta o restes de menjar, o insectes i altres ectoparàsits, els lèmurs s'empolainen amb la llengua i es raspallen amb la pinta dental.[12][100] Malgrat aquestes diferències tècniques, els lèmurs s'empolainen amb la mateixa freqüència i pels mateixos motius que els antropoïdeus.[100]
Patrons d'activitat
[modifica]El seu ritme biològic pot variar des d'un comportament nocturn en els lèmurs més petits a diürn en la majoria dels més grans. La diürnalitat no es dona en cap altre prosimi.[27] La catemeralitat (activitat tant de dia com de nit) es dona en alguns lèmurs més grans; pocs altres primats presenten aquest model d'activitat[101] quan canvien les condicions ambientals.[12] Els lèmurs catemerals que s'han estudiat amb més detall són els lèmurs vertaders (gènere Eulemur).[39][102] Tot i que el lèmur mangosta és el millor documentat, totes les espècies estudiades d'aquest gènere presenten un cert grau de comportament catemeral,[67] malgrat que la seva activitat nocturna sovint està restringida per la disponibilitat de llum i les fases lunars.[13] Aquest tipus de comportament fou documentat per primera vegada a la dècada del 1960 en espècies de lèmurs vertaders, així com en altres espècies de lemúrids, com ara els lèmurs de collar i els lèmurs del bambú. Inicialment descrit com a «crepuscular» (actiu a l'alba i el vespre), l'antropòleg Ian Tattersall dugué a terme investigacions addicionals i encunyà el nou terme «catemeral»,[101] tot i que altres científics que no són antropòlegs prefereixen els termes «circadiari» o «nictimeral».[13]
Per tal de conservar energia i aigua al seu medi tan estacional,[84][103] els lèmurs ratolí i els lèmurs nans han desenvolupat cicles estacionals de dormància durant els quals el seu ritme metabòlic i la seva temperatura corporal baixen. Són els únics primats coneguts que ho fan.[84] Acumulen reserves de greix als membres inferiors i la base de la cua abans de l'àrid hivern, quan escassegen el menjar i l'aigua,[28][85] i poden presentar períodes perllongats de letargia diària durant l'estació seca. Aquesta letargia consisteix en menys de 24 hores de dormància, mentre que la mitjana de les letargies perllongades assoleix les dues setmanes de durada i indica que s'acosta la hibernació.[84] Els lèmurs ratolí han sigut observats en estat de letargia durant dies, però se sap que els lèmurs nan passen entre sis i vuit mesos a l'any hibernant,[27][28][86] especialment a la costa occidental de Madagascar.[103]
Els lèmurs nans són els únics primats coneguts que hibernen durant períodes llargs.[84][104] A diferència d'altres mamífers hibernants de regions temperades, que periòdicament es desperten durant uns dies, els lèmurs nans tenen una hibernació contínua de maig a setembre. Abans i després d'aquesta hibernació profunda, hi ha dos mesos de transició (abril i octubre) durant els quals busquen aliment de manera limitada per reduir la despesa de reserves de greix.[103] A diferència de qualsevol altre mamífer que hiberni, la temperatura corporal dels lèmurs nans hibernants fluctua amb la temperatura ambient, en lloc de mantenir-se baixa i estable.[28][86][103]
Altres lèmurs que no passen períodes de dormància conserven energia seleccionant microhàbitats termoregulats (com ara forats als arbres), compartint el niu i reduint la superfície corporal exposada, en posar-se corbats i arraulir-se en grup. Sovint es veuen els lèmurs de cua anellada, lèmurs de collar i sifaques prenent el sol, cosa que els serveix per escalfar-se sense haver de recórrer a la calor corporal generada pel metabolisme.[84]
Locomoció
[modifica]
El comportament locomotor dels lèmurs, tant vivents com extints, és molt variat i la seva diversitat supera la dels primats antropoïdeus.[36] Les postures i els comportaments locomotors inclouen el salt i l'aferrament vertical,[N 5] propi dels lèmurs del bambú i dels indris;[36][56] el desplaçament quadrúpede arbori lent (com els loris), que en el seu dia utilitzava el mesopropitec;[106] el desplaçament quadrúpede arbori ràpid, com en el cas dels lèmurs marrons i els lèmurs rufos;[36][107] la locomoció cuadrúpeda parcialment terrestre, observada en el lèmur de cua anellada; la locomoció quadrúpeda fonamentalment terrestre, desenvolupada antany pels lèmurs mico, com ata Hadropithecus;[36] i el desplaçament en suspensió, similar al dels peresosos, que mostraven en el seu dia els lèmurs peresós, com ara Palaeopropithecus.[12][36] S'ha observat que el lèmur gris del llac Alaotra és un bon nedador.[12] A vegades, aquests sistemes de locomoció s'agrupen en dos tipus, els d'agafament i salt vertical i els quadrúpedes arboris (a vegades també terrestres).[64]
El poderós salt dels índrids està ben documentat i és molt popular entre els ecoturistes que visiten Madagascar.[108] Utilitzant les seves potes llargues i fortes es catapulten cap amunt i cauen a terra o en un arbre proper n una postura erecta i s'aferren amb força al tronc amb els peus i les mans.[14] Els indris poden desplaçar-se amb rapidesa d'un tronc d'arbre a l'altre fent salts de fins a deu metres,[14][60] una capacitat coneguda com a ricochetal leaping ('salt de rebots').[74] El sifaca de Verreaux és capaç de realitzar aquest tipus de salts als boscos espinosos del sud de Madagascar; es desconeix com pot evitar que les mans li quedin traspassades als troncs coberts d'espines de plantes grans com ara Alluaudia.[14] Quan la separació entre els arbres és massa gran, el sifaca baixa a terra i cobreix distàncies de més de cent metres mentre roman dempeus i saltant lateralment, agitant el pit de dalt a baix per guanyar alçada i deixant els braços oberts, presumiblement per mantenir l'equilibri.[14][60] Aquest moviment es descriu a vegades com a «salt de ball».[14]
Comunicació
[modifica]Els lèmurs poden comunicar-se mitjançant sons i a través de la visió i l'olfacte. El lèmur de cua anellada, per exemple, utilitza models complexos tot i que molt estereotipats, com ara marcatge territorial i vocalitzacions.[83] Els senyals visuals són probablement els que menys utilitzen els lèmurs, car manquen de molts dels músculs utilitzats per altres primats per a les expressions facials.[80] A causa la seva pobra visió, les postures de cos sencer són generalment més identificables. Tanmateix, s'ha comprovat que el lèmur de cua anellada utilitza diferents expressions facials davant de diverses situacions, com una mirada fixa amenaçadora, tirar enrere els llavi en senyal de submissió o girar cap enrere les orelles i eixamplar les fosses nasals durant el marcatge territorial.[83] També s'ha vist aquesta espècie utilitzant badalls com a senyal d'amenaça.[109][110] La seva cua anellada també comunica distanciament, adverteix grups veïns i ajuda a localitzar membres del seu grup.[83] Se sap que els sifaques utilitzen un gest de joc amb la boca oberta,[111] així com una ganyota submisa ensenyant les dents, utilitzada en interaccions agonístiques.[61]

L'olfacte és particularment important pels lèmurs,[12] tret de l'indri, que manca de la majoria de glàndules odoríferes comunes als altres lèmurs, a part de tenir el cervell amb una regió olfactòria molt reduïda.[74] A través de l'olfacte poden transmetre informació sobre edat, sexe o estatus reproductiu, així com demarcar els límits d'un territori. És el sentit més útil perquè es comuniquin animals que rarament es troben els uns amb els altres.[46] els lèmurs petits i nocturns marquen territori amb orina, mentre que les espècies més grosses i diürnes utilitzen glàndules odoríferes localitzades a diferents parts del cos. El lèmur de cua anellada participa en «lluites pudents», fregant-se la cua amb les glàndules d'olor dels canells per després fuetejar-hi els mascles rivals. Alguns lèmurs defequen en àrees específiques (tret conegut a vegades com a «comportament de latrina»). Encara que molts animals presenten aquest comportament, és un tret rar entre els primats. Aquesta conducta a l'hora de defecar pot representar un marcatge territorial i una ajuda en la senyalització interespecífica.[13]
Comparats amb els altres mamífers, els primats en general són molt vocals i els lèmurs no en són una excepció.[13] Algunes espècies de lèmur tenen extensos repertoris vocals, com el lèmur de cua anellada i els lèmurs de collar.[83][112] Algunes de les crides més comunes entre els lèmurs són les d'alarma de depredadors. Els lèmurs no només reaccionen davant de crides d'alarma de les seves pròpies espècies, sinó també davant de les crides d'alarma d'altres espècies o aus no depredadores. El lèmur de cua anellada i alguns altres lèmurs tenen diferents crides i reaccions davant de tipus específics de depredadors.[36] Amb les crides d'aparellament, s'ha comprovat que els lèmurs ratolí que no poden discernir-se visualment responen amb més força a les crides de la seva pròpia espècie, especialment quan són les crides d'altres lèmurs ratolí que es troben habitualment al seu territori.[113] Les crides dels lèmurs poden ser molt forts i arribar a grans distàncies. Els lèmurs de collar utilitzen crides que es poden sentir fins a un quilòmetre de distància durant un dia clar i en calma.[112] El lèmur més sorollós és l'indri, les crides del qual es poden sentir fins a dos quilòmetres o més de distància,[47][59] per poder comunicar de manera més eficaç els límits del seu territori, que tenen una extensió d'entre 34 i 40 hectàrees.[74] Tant els lèmurs de collar com els indris presenten un comportament de crides contagioses, en el qual un individu o grup comença una crida forta i altres de la mateixa àrea se li uneixen.[59][112] El cant de l'indri pot durar des de 45 segons fins a més de 3 minuts. Aquests animals tendeixen a coordinar-se per formar un duo estable comparable al dels gibons.[59][58]
La comunicació tàctil s'utilitza fonamentalment en forma d'empolainament social, tot i que els lèmurs de cua anellada també es reuneixen en grups per dormir junts (en un ordre determinat pel seu rang), allargant la mà i tocant alguns membres adjacents del grup, o donant manotades als altres. En aquesta espècie, allargar la mà i tocar altres individus s'ha vist com a possible conducta de submissió, realitzada per animals més joves o submisos envers els membres més vells i més dominants del grup. Tanmateix, l'empolainament social sembla donar-se més sovint entre individus de major posició, un tret compartit amb altres espècies de primats.[114] A diferència d'allò que passa en els primats antropoïdeus, l'empolainament en els lèmurs sembla més íntim i sovint és recíproc. Els antropoïdeus, d'altra banda, utilitzen l'empolainament social per gestionar interaccions agonístiques.[115] Se sap que el lèmur de cua anellada és molt tàctil i dedica entre un 5 i un 11% del seu temps a l'empolainament.[114]
| Mostra de vocalitzacions de lèmurs | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|

Problemes de reproducció? Vegeu l'ajuda
|
Problemes de reproducció? Vegeu l'ajuda
| ||||||||||||||||
Defensa contra depredadors
[modifica]Tots els lèmurs pateixen algun tipus de pressió per depredadors.[116] Entre les mesures de defensa comunitària contra la depredació es troba la utilització de crides d'alarma i l'atac o assetjament col·lectiu (mobbing) contra l'agressor,[117] sobretot entre els lèmurs diürns.[36] Segons un estudi cinemàtic, les capacitats de salt dels lèmurs podrien haver evolucionat per a la defensa contra depredadors, més que per desplaçar-se.[118] Els lèmurs nocturns són difícils de veure i rastrejar de nit. Encara dificulta més la seva visibilitat que busquin aliment en solitari. També intenten evitar els depredadors utilitzant posicions encobertes per dormir, com ara nius, forats als arbres o vegetació densa,[36] i canviant entre diverses localitzacions.[28] Fins i tot la letargia i els estats d'hibernació dels quirogalèids podrien ser deguts en part al fet que pateixen nivells elevats de depredació.[116] Les cries es mantenen protegides durant la cerca d'aliment, sia deixant-les al niu o amagant-les en un lloc ocult, on la cria roman immòbil mentre els pares són fora.[36]
Els lèmurs diürns són visibles durant el dia, per la qual cosa viuen en grups, on un major nombre d'ulls i orelles ajuda a descobrir possibles depredadors. Els lèmurs diürns utilitzen i responen a les crides d'alarma, fins i tot les d'altres espècies de lèmur i les d'aus no depredadores. El lèmur de cua anellada té diferents crides i reaccions davant de diferents depredadors, com les aus de presa, els mamífers o les serps.[36] Alguns, com ara l'indri, utilitzen una estratègia de cripsi per camuflar-se. Sovint se'ls sent, però són difícils de veure entre els arbres a causa de la llum clapada (mescla de llum i ombra) dels boscos, cosa que els ha donat la reputació de ser «fantasmes del bosc».[74]
Reproducció
[modifica]A part de l'ai-ai i el lèmur gris del llac Alaotra, els lèmurs es reprodueixen estacionalment,[12][39] amb temporades d'aparellament i naixement molt curtes a causa de la influència de la disponibilitat molt estacional de recursos al seu entorn. Les èpoques de zel duren generalment menys de tres setmanes cada any,[36] i l'obertura vaginal de les femelles només es produeix durant uns dies o unes hores del període més receptiu de l'estre.[79] Aquests estrets marges per a la reproducció i la disponibilitat de recursos semblen estar relacionats amb la gestació curta, la ràpida maduració i el baix metabolisme basal, així com un elevat cost energètic per a les femelles a l'hora de reproduir-se. També pot estar relacionat amb la taxa de mortalitat relativament alta que es dona entre les femelles adultes i una major proporció de mascles adults en algunes poblacions de lèmurs (ambdós trets són rars entre els primats). En el cas de l'ai-ai i el lèmur gris del llac Alaotra, els naixements es produeixen al llarg d'un període de sis mesos.[12]
Encaixen les temporades d'aparellament i naixement de manera que els períodes de deslletament estiguin sincronitzats per coincidir amb el moment de major disponibilitat de menjar.[79][88] El deslletament es produeix una mica abans o després de l'erupció dels seus primers queixals permanents.[30] Els lèmurs ratolí són capaços d'ajustar el seu cicle de cria complet a l'estació humida, mentre que els lèmurs més grans, com els sifaques, han de lactar dos mesos durant la temporada seca.[88] S'ha comprovat que la supervivència infantil en algunes espècies, com el sifaca de Milne-Edwards, es veu afectada directament tant per les condicions ambientals com pel rang, l'edat i la salut de la seva mare. La temporada de cria també és afectada per la situació geogràfica; per exemple, els lèmurs ratolí pareixen entre setembre i octubre al seu hàbitat natal de l'hemisferi sud, però els exemplars en captivitat a l'hemisferi nord ho fan de maig a juny.[79]
Els factors odorífers tenen una gran importància en la reproducció dels lèmurs. L'activitat de marcatge territorial s'intensifica durant l'època de zel, i les feromones poden coordinar el moment reproductiu per a les femelles que entren en estre.[79] L'aparellament pot ser monògam o promiscu, tant en els mascles com en les femelles, i l'acoblament pot incloure individus aliens al grup.[12][36] Entre els lèmurs monògams hi ha el lèmur de panxa vermella i el lèmur mangosta, tot i que aquest últim ha sigut observat aparellant-se fora del seu vincle de parella.[36] La monogàmia és més comuna entre les espècies nocturnes, tot i que s'hi donen mostres de competicions de lluita, supressió sexual de subordinats o competència entre mascles que eviten enfrontaments directes.[30] En el cas dels lèmurs ratolí, els mascles utilitzen el tap de copulació, un augment del desenvolupament dels testicles durant l'època de zel i un increment en el dimorfisme de mida.[119]

El període de gestació varia segons l'espècie i va des de les nou setmanes en els lèmurs ratolí a entre nou i deu en els lèmurs nans o fins i tot divuit o vint-i-quatre setmanes en altres lèmurs.[79] Les espècies més petites i nocturnes, com les del gènere Mirza, els lèmurs ratolí i els lèmurs nans, solen donar a llum més d'una cria, mentre que els lèmurs més grans i nocturns, com els lèmurs forcats, els lèmurs mostela i l'ai-ai en general tenen un descendent.[27] Els lèmurs nans i els lèmurs ratolí tenen fins a quatre descendents, però en ambdues espècies la mitjana és de només dos. Els lèmurs de collar són els únics lèmurs grans i diürns que pareixen de forma habitual dues o tres cries. Totes les altres espècies solen donar a llum una sola cria. Els parts múltiples en els lèmurs són normalment dizigòtics i solen produir-se en un de cada cinc o sis naixements en espècies com el lèmur de cua anellada o algunes del gènere Eulemur.[79]
Després del naixement, els lèmurs solen amagar les cries o endur-se-les mentre busquen aliment. Quan porten les cries, aquestes s'aferren a la pell de la mare o són portades agafades pel coll amb la boca. En algunes espècies, com els lèmurs grisos, els petits són portats amb la boca fins que són capaços d'aferrar-se a la pell de la seva mare.[120] Entre les espècies que carreguen la seva progènie s'inclouen les espècies nocturnes (com els lèmurs ratolí, els lèmurs mostela i els lèmurs nans), els lèmurs grisos i els lèmurs de collar.[27][120] En el cas dels lèmurs de collar, els petits són altricials i les mares els construeixen nius, igual que les espècies de lèmur més petites i nocturnes.[12] Els indris llanosos són una excepció entre els lèmurs nocturns perquè viuen en grups familiars cohesionats i s'enduen la seva única cria en lloc de deixar-la a l'àrea de cria.[56][57] S'ha descrit la cria o cura al·loparental (assistència per individus diferents dels progenitors a l'hora de cuidar de la prole, sia de manera individual o pel grup) en totes les famílies de lèmurs tret dels lèmurs mostela i l'ai-ai. També se sap que l'al·loalletament (cessió de llet a cries d'altres mares) es dona en diversos grups de lèmurs.[121] S'ha comprovat que fins i tot els mascles poden tenir cura de les cries en algunes espècies, com el lèmur de panxa vermella, el lèmur mangosta,[67] el lèmur gris oriental, el sifaca sedós,[121] el lèmur nan de cua gruixuda[122] o els lèmurs de collar.[123]
Una altra de les característiques que diferencia la major part dels lèmurs dels primats antropoïdeus és la seva longevitat i la seva alta taxa de mortalitat infantil.[88] Molts lèmurs, incloent-hi el lèmur de cua anellada, s'han adaptat a un ambient altament estacional, cosa que n'ha afectat l'índex de natalitat, la maduresa i la taxa d'aparellament (selecció r). Això els ajuda a recuperar-se ràpidament d'una davallada demogràfica.[83] Poden viure el doble en captivitat que a la natura perquè es beneficien d'una nutrició consistent que satisfà les seves necessitats alimentàries, dels progressos mèdics i de la millora en la compressió dels seus requisits d'allotjament. El 1960 es creia que els lèmurs podrien viure entre 23 i 25 anys, però actualment se sap que les espècies més grosses poden viure més de 30 anys sense mostrar senyals d'envelliment (senescència) i mantenint la seva capacitat reproductiva.[79]
Habilitats cognoscitives i ús d'eines
[modifica]Tradicionalment s'ha considerat els lèmurs menys intel·ligents que els primats antropoïdeus[124] i s'ha descrit els micos i hominoïdeus com a més astuts, hàbils i entabanadors.[14] Moltes espècies de lèmur, com els sifaques i el lèmur de cua anellada, han donat pitjors resultats en proves dissenyades per a micos, però tenen el mateix rendiment en d'altres.[14][100] Aquestes comparacions podrien ser injustes, car els lèmurs prefereixen manipular els objectes amb la boca més que amb les mans i només s'interessen per objectes quan estan en captivitat.[100] De fet, diversos estudis han demostrat que els lèmurs tenen el mateix nivell d'intel·ligència tècnica que la resta de primats.[125] No s'han vist lèmurs utilitzant eines a la natura, tot i que en captivitat el lèmur bru i el lèmur de cua anellada han demostrat ser capaços d'entendre i utilitzar eines.[13]
El cervell d'algunes espècies és relativament gran. L'hadropitec, un gènere extint, tenia la mida corporal d'un babuí mascle gros i un cervell de mida comparable, cosa que en fa el cervell més gros en relació a la massa corporal de tots els prosimis.[126] L'ai-ai també té una proporció cervell-massa corporal gran, fet que pot indicar un nivell més alt d'intel·ligència.[39] Tanmateix, malgrat tenir un instrument en forma del seu llarg i prim dit mitger, que utilitza per capturar larves d'insecte, l'ai-ai no ha tingut un rendiment gaire bo en les proves d'utilització d'eines.[13]
Ecologia
[modifica]Madagascar no només té dues zones climàtiques radicalment diferents (les selves tropicals de l'est i les regions seques de l'oest),[12] sinó que també pateix des de profundes sequeres fins a inundacions provocades per ciclons.[127] Aquests reptes climàtics i geogràfics, juntament amb uns sòls pobres i de baixa productivitat vegetal, la gran varietat i complexitat dels seus ecosistemes i la manca d'arbres que donin fruit amb regularitat (com les figueres) han portat els lèmurs a evolucionar cap a una enorme diversitat morfològica i de comportament.[11][12][30][88] La seva supervivència ha requerit la capacitat de suportar extrems constants, en lloc d'una estabilitat o regularitat anual.[127]


Els lèmurs ocupen o han ocupat els nínxols ecològics generalment omplerts per micos, esquirols, picots i ungulats pasturadors.[14] Atesa la seva diversitat d'adaptacions a nínxols ecològics específics, la selecció d'hàbitat entre les famílies i alguns gèneres de lèmurs sovint és molt diferent, cosa que minimitza la competència.[12] Fins a cinc espècies de lèmurs nocturns dels boscos més estacionals de l'oest poden coexistir durant l'estació humida gràcies a la gran abundància de menjar. Tanmateix, per suportar l'extrema estació seca de l'illa, tres d'aquestes cinc espècies utilitzen diferents models alimentaris (amb els seus trets fisiològics subjacents) per poder coexistir: els lèmurs forcats s'alimenten de la goma dels arbres, els lèmurs mostela de les fulles i els del gènere Mirza a vegades s'alimenten de secrecions d'insectes. Les altres dues espècies, el lèmur ratolí gris i el lèmur nan de cua gruixuda, eviten la competència reduint l'activitat: el lèmur ratolí gris es val de temporades de letargia, mentre que el lèmur nan de cua gruixuda hiberna completament.[27] D'igual manera, a la costa oriental gèneres sencers se centren en un menjar específic per evitar una excessiva superposició de nínxols; els lèmurs bruns i els lèmurs de collar són frugívors, els índrids són folívors i els lèmurs grisos s'especialitzen en el bambú i altres herbes. Una vegada més, les diferències alimentàries estacionals, així com les subtils diferències en les preferències de substrat, els estrats forestals utilitzats, el cicle d'activitat i l'organització social permeten la coexistència de diferents espècies de lèmurs, tot i que en aquest cas les espècies estan més estretament relacionades i tinguin nínxols similars.[12] Un bon exemple és la divisió de recursos entre tres espècies de lèmurs grisos que viuen molt properes en petites àrees poblades d'arbres, el lèmur daurat, el lèmur de nas ample i el lèmur gris oriental: cadascun utilitza espècies diferents de bambú, parts diferents de la planta o capes diferents del bosc.[14][51] El contingut de nutrients i toxines (com ara cianur) ajuda a regular la selecció de menjar,[12] tot i que se sap que les preferències estacionals d'aliment també acompleixen un paper en la selecció.[51]
Tenen règims alimentaris variats segons les espècies: n'hi ha de nectarívors, folívors, frugívors o omnívors, amb espècies molt adaptables i d'altres amb una dieta especialitzada, com ara exsudacions de plantes o bambú.[68] En alguns casos, els patrons alimentaris dels lèmurs beneficien la vida vegetal local. Quan s'alimenten de nèctar poden actuar com a pol·linitzadors, mentre que les parts funcionals de la flor no queden danyades.[87] De fet, diverses plantes amb flor malgaixes no relacionades mostren trets de pol·linització específics dels lèmurs i diversos estudis indiquen que algunes espècies diürnes, com ara el lèmur de panxa vermella i els lèmurs de collar, actuen com a pol·linitzadors primaris.[12] Dos exemples d'espècies de plantes que es valen dels lèmurs per a la pol·linització són l'arbre del viatger[54] i una espècie de liana semblant a un llegum, Strongylodon cravieniae.[12] La dispersió de les llavors és un altre dels serveis facilitats pels lèmurs; després de passar pel ventre del lèmur, les llavors de les plantes enfiladisses i els arbres pateixen una mortalitat inferior i germinen més ràpidament.[88] El comportament de defecar en algunes àrees específiques que tenen alguns lèmurs pot ajudar a millorar la qualitat del sòl i facilitar la dispersió de llavors.[13] A causa de la seva importància per al manteniment d'un bosc sa, els lèmurs frugívors poden qualificar-se com a mutualistes clau.[88]
Tots els lèmurs, particularment les espècies més petites, són víctimes de la depredació,[27][116] i constitueixen una presa important per a diversos depredadors.[119] Els humans són els depredadors més significatius dels lèmurs diürns, malgrat que alguns tabús prohibeixen caçar o alimentar-se de certes espècies de lèmur.[12] Entre els seus depredadors hi ha euplèrids nadius com ara la civeta de Madagascar i la fossa, gats salvatges, gossos domèstics, serps, rapinyaires diürns i nocturns i cocodrils. Àguiles gegants extintes, com ara Stephanoaetus mahery, i una o dues espècies del gènere Aquila, així com la fossa de les cavernes, antany també s'alimentaren de lèmurs, potser fins i tot dels lèmurs gegants, que s'extingiren recentment, o les seves cries subadultes.[27][116] L'existència d'aquests gegants extints suggereix que les interaccions entre presa i depredador que implicaven els lèmurs eren més complexes que les actuals.[12] Avui en dia, la mida dels depredadors limita els mussols als lèmurs més petits, en general de 100 g o menys, mentre que els lèmurs de mida més gran cauen víctimes de rapinyaires diürns més grans, com ara l'arpella esparverenca de Madagascar i l'aligot de Madagascar.[116]
Recerca
[modifica]Les semblances dels lèmurs amb els simis, com la dieta i l'organització social, juntament amb els seus propis trets distintius, han fet que els lèmurs hagin sigut el grup de mamífers de Madagascar més estudiat.[12][63] Les investigacions sovint se centren en la relació entre la seva ecologia i la seva organització social, però també en el comportament i la morfofisiologia (estudi de l'anatomia en relació a la seva funció).[12] Els estudis sobre les característiques del seu cicle vital, el seu comportament i la seva ecologia ajuden a entendre l'evolució dels primats, car es creu que comparteixen trets amb els primats ancestrals. D'altra banda, per la seva proximitat filogenètica als humans i igual que altres primats, els lèmurs són subjectes d'experiments en l'estudi de malalties de gran incidència i mortalitat entre els humans, com ara la malaltia d'Alzheimer[128][129] o la sida.[130]
Han sigut l'eix central de sèries monogràfiques, plans d'acció, guies de camp i obres clàssiques en etologia.[63] Tanmateix, fins ara s'han estudiat poques espècies a fons i gran part de la recerca ha sigut preliminar i s'ha restringit a una sola localització.[12] Fa poc que s'han començat a publicar nombrosos articles científics que expliquen aspectes bàsics del comportament i l'ecologia d'espècies poc conegudes. Els estudis de camp han donat una nova percepció d'aquests primats en camps com ara la dinàmica de poblacions i l'ecologia evolutiva de la majoria de gèneres i moltes espècies.[63] Les investigacions a llarg termini centrades en individus identificats estan a la seva infància i només han començat amb unes poques poblacions. Tanmateix, les oportunitats d'aprenentatge disminueixen a causa de la destrucció del seu hàbitat i altres factors que amenacen l'existència de les poblacions de lèmurs arreu de l'illa.[12]

Els lèmurs ja foren mencionats pels marins als quaderns de bitàcola dels seus viatges el 1608 i el 1658, i almenys set espècies foren descrites en detall pel governador francès de Madagascar Étienne de Flacourt, que també pot ser que hagi sigut l'únic occidental que veié i feu una crònica sobre un lèmur gegant (actualment extint) que anomenà tretretretre.[131][N 6] Cap al 1703, els comerciants i marins començaren a dur lèmurs a Europa. James Petiver, un apotecari de Londres, descrigué i il·lustrà el lèmur mangosta. A partir del 1751, el naturalista londinenc George Edwards començà a descriure i il·lustrar diverses espècies de lèmurs, algunes de les quals foren incloses per Carl von Linné en diverses edicions del seu Systema Naturae. A les dècades del 1760 i 1770, els naturalistes francesos Georges Louis Leclerc i Louis Jean-Marie Daubenton començaren a descriure l'anatomia de diverses espècies de lèmurs. El primer naturalista de camp a confeccionar informes sobre els lèmurs fou Philibert Commerson el 1771, tot i que fou Pierre Sonnerat qui registrà la major varietat d'espècies durant els seus viatges.[127][133]
Durant el segle xix hi hagué una explosió de nous noms i descripcions de lèmurs, que posteriorment trigaren dècades a ser classificats. Durant aquest temps, els colectores professionals recolliren espècimens per a museus, «cases de feres» i gabinets de curiositats. Alguns dels principals col·leccionistes foren Johannes Hildebrandt i Charles Immanuel Forsyth Major. A partir d'aquestes col·leccions, així com de les creixents observacions de lèmurs als seus hàbitats naturals, sistemàtics com ara Albert Günther i John Edward Gray continuaren aportant nous noms per a les espècies de lèmur. Tanmateix, la contribució més notable d'aquest segle fou possiblement l'obra d'Alfred Grandidier, un naturalista i explorador que es dedicà a l'estudi de la història natural de Madagascar i la seva població local. Amb l'ajuda d'Alphonse Milne-Edwards, la majoria dels lèmurs diürns foren il·lustrats en aquell temps. Tanmateix, la nomenclatura taxonòmica dels lèmurs assolí la seva forma moderna a les dècades del 1920 i del 1930, quan fou estandarditzada pel zoòleg alemany Ernst Schwarz el 1931.[127][133]
Tot i que la seva taxonomia havia experimentat un important desenvolupament, no fou fins a les dècades del 1950 i del 1960 que l'estudi in situ del seu comportament i la seva ecologia començà a florir. Jean-Jacques Petter i Arlette Petter-Rousseaux recorregueren Madagascar el 1956 i el 1957, fent el reconeixement d'un gran nombre d'espècies de lèmurs i importants observacions sobre els seus grups socials i la seva reproducció. El 1960, any de la independència de Madagascar, David Attenborough presentà als lèmurs a Occident a través d'una pel·lícula comercial. Sota la direcció de John Buettner-Janusch, que fundaria el Duke Lemur Center el 1966, Alison Jolly viatjà a Madagascar el 1962 per estudiar la dieta i el comportament social del lèmur de cua anellada i el sifaca de Verreaux a la reserva privada de Berenty. Jolly i els Petter assentaren les bases d'una nova era d'interès per l'ecologia i el comportament dels lèmurs i aviat foren secundats per antropòlegs com ara Alison Richard, Robert Sussman i Ian Tattersall. Després de la pausa provocada per la confusió política de mitjans de la dècada del 1970 i la revolució de Madagascar, els estudis de camp continuaren a la dècada del 1980, gràcies en part a la renovada participació del Duke Lemur Center sota la direcció d'Elwyn Simons i els esforços ecologistes de Patricia Wright.[12][127][133] En les dècades posteriors s'ha fet un gran salt en l'estudi dels lèmurs i s'ha descobert un gran nombre de noves espècies.[5]
Els estudis fora de Madagascar també resulten atractius per als investigadors que busquen resposta a qüestions que són difícils de posar a prova a la natura. Per exemple, els estudis per a la seqüenciació completa del genoma de determinats lèmurs ajudaran els investigadors a entendre quins trets genètics diferencien els primats dels altres mamífers i contribuiran a comprendre quins factors genòmics diferencien els humans dels altres primats.[31] Una de les principals instal·lacions d'investigació de lèmurs és el Duke Lemur Center, situat a Durham (Carolina del Nord). Aquest centre associat a la Universitat Duke té la població de lèmurs en captivitat més nombrosa que hi ha fora de Madagascar, que serveix per dur a terme recerca no invasiva i un programa de reproducció en captivitat. [134] Al centre s'han dut a terme importants projectes d'investigació, com ara estudis de les seves vocalitzacions,[135] el seu comportament locomotor,[136] la cinemàtica del bipedisme,[137] els efectes de la seva complexitat social sobre el raonament transitiu[N 7][138] i estudis sobre cognició que analitzen la capacitat d'un lèmur d'organitzar i recuperar seqüències de la seva memòria.[139] Altres instal·lacions, com ara la Lemur Conservation Foundation, localitzada prop de Myakka City (Florida), també han dut a terme projectes d'investigació, com un estudi sobre la capacitat dels lèmurs de seleccionar eines basant-se preferentment en les seves qualitats funcionals.[140]
Estat de conservació
[modifica]

Els lèmurs estan amenaçats per un gran nombre de problemes ambientals, com la desforestació, la caça per al consum de la seva carn, la captura d'espècimens vius per al comerç d'animals de companyia exòtics i el canvi climàtic.[141][88] Totes les espècies de lèmur estan incloent-hi a l'Apèndix I de la CITES, que prohibeix el comerç d'espècimens o de les seves parts si no estan destinats a investigacions científiques.[142][143] Des del 2005, la Unió Internacional per a la Conservació de la Natura (UICN) inclogué en la seva Llista Vermella d'Espècies Amenaçades un 16% dels lèmurs com a espècies en perill crític, un 23% com a espècies en perill, un 25% com a espècies vulnerables i un 28% com a espècies amb dades insuficients. Tan sols un 8% tenia la consideració d'espècie en risc mínim.[134] Durant els cinc anys següents se n'identificaren almenys 28 noves espècies, però encara no s'ha establert l'estat de conservació de cap d'elles.[42] És probable que moltes d'elles s'acabin considerant com amenaçades, car les espècies descrites fa poc temps generalment es concentren en regions petites.[144] Considerant el progrés continu de la destrucció del seu hàbitat, algunes espècies encara no descobertes podrien extingir-se abans de ser identificades.[63] Des de l'arribada dels humans a l'illa, fa aproximadament 2.000 anys, tots els vertebrats endèmics de Madagascar de més de 10 kg de pes han desaparegut,[35] incloent-hi 17 espècies, 8 gèneres i 3 famílies de lèmurs.[34][37] La Comissió de Supervivència d'Espècies de la UICN (IUCN/SSC), la Societat Primatològica Internacional (IPS) i Conservation International (CI) han inclòs almenys cinc lèmurs en la seva publicació biennal Els 25 primats més amenaçats del món. L'informe corresponent als anys 2012-2014 n'inclou sis: Eulemur flavifrons, Lepilemur septentrionalis, Propithecus candidus, Microcebus berthae, Indri indri i Varecia rubra.[145] El 2012, una avaluació del Grup d'Experts en Primats de la UICN conclogué que un 91% de les 103 espècies de lèmurs conegudes aleshores estaven amenaçades, cosa que en fa el grup de mamífers en major risc d'extinció.[146][147] El 2013, la UICN reiterà la seva preocupació i avisà que el 90% de les espècies de lèmurs podrien extingir-se en dels propers 20-25 anys llevat que s'implementi un pla de conservació de tres anys per ajudar les comunitats locals, que costaria 7 milions de dòlars.[148][149]
Madagascar és un dels països més pobres del món,[150][151] amb una taxa de creixement poblacional del 2,5% i gairebé el 70% de la població vivint en la pobresa.[36][150] El país té un deute extern elevat i disposa de pocs recursos.[151] Aquestes qüestions socioeconòmiques fan que resulti complicat aportar recursos dedicats a la conservació de la naturalesa, tot i que l'illa de Madagascar ha sigut declarada per la Comissió de Supervivència d'Espècies de la UICN com a regió de primats crítica des de fa més de vint anys.[144] A causa de la seva petita superfície (587.045 km²) en comparació amb altres regions d'alta biodiversitat i el seu elevat nivell d'endemismes, el país és considerat un dels punts calents de biodiversitat (més importants del món, amb la conservació dels lèmurs qualificada d'objectiu d'alta prioritat.[134][144] Malgrat l'èmfasi posada en els esforços de conservació, no hi ha cap indici que les extincions que començaren amb l'arribada dels humans hagin tocat a la seva fi.[35]
Amenaces a la natura
[modifica]La major amenaça a la qual s'enfronten les poblacions de lèmurs és la destrucció i degradació del seu hàbitat.[36][142] La desforestació sovint és provocada per les necessitats de subsistència local, com l'agricultura de tala i crema (tavy, en malgaix), la creació de zones de pastura per al bestiar per mitjà de la incineració, la provisió legal i il·legal de fusta per a llenya o producció de carbó vegetal, la mineria comercial i la tala il·legal de fustes nobles per a mercats exteriors.[36][141] Després de segles d'ús insostenible, així com la ràpida i progressiva intensificació de la destrucció forestal des del 1950,[134] queden menys de 60.000 km² (el 10% de la superfície terrestre) poblats d'arbres a Madagascar. Tan sols 17.000 km² de l'àrea terrestre (3%) de l'illa estan protegits i, a causa d'unes condicions econòmiques extremes i la inestabilitat política, la majoria de les àrees protegides són administrades i defensades de forma ineficaç.[141][144] Algunes àrees protegides foren deixades de banda perquè estaven salvaguardades de manera natural per la seva situació aïllada, remota i sovint escarpada. altres àrees, com els boscos secs i els boscos espinosos de l'oest i sud de l'illa, reben poca protecció i estan en gran perill de ser destruïdes.[36]
Algunes espècies poden estar en risc d'extinció fins i tot sense una desforestació completa, com els lèmurs de collar, que són molt sensibles a la pertorbació del seu hàbitat.[63] Si s'eliminen els grans arbres fruiters, el bosc pot mantenir menys individus d'una espècie i el seu èxit reproductiu pot veure's afectat durant anys.[88] Les poblacions petites poden persistir a fragments forestals aïllats durant 20 o 40 anys gràcies als seus perllongats períodes generacionals, però a llarg termini aquestes poblacions poden no ser viables.[152] Les poblacions petites i aïllades també es podrien extingir a causa de catàstrofes naturals o epidèmies. Dues malalties que són letals per als lèmurs i podrien afectar greument poblacions aïllades són la toxoplasmosi, propagada per gats salvatges, i el virus de l'herpes simple, transmès pels humans.[153]

El canvi climàtic i les catàstrofes naturals relacionades amb el clima també n'amenacen la supervivència. Durant els últims 1.000 anys, les regions occidentals i muntanyoses han esdevingut considerablement més seques, però en les últimes dècades les sequeres greus s'han tornat molt més freqüents. Hi ha indicis que la desforestació i la fragmentació forestal acceleren aquesta dessecació gradual.[88] Els efectes de la sequera s'aprecien fins i tot a les selves tropicals. Com a conseqüència de la disminució de les precipitacions anuals, els arbres més grans que formen el cobricel de les zones boscoses pateixen un augment de la seva mortalitat, una menor eficàcia dels seus fruits i una disminució de la producció de fulles noves, que són les que prefereixen els lèmurs folívors. Els ciclons poden desfullar algunes àrees, fer caure arbres del cobricel i provocar despreniments de terra i inundacions. Això pot deixar algunes poblacions de lèmurs sense fruita o fulles fins a la primavera següent i obligar-les a subsistir a còpia d'aliments de crisi, com ara epífits.[154]
Els lèmurs són caçats com a aliment per la població nadiua, tant per a la caça de subsistència com amb fins comercials per aprovisionar de carn de luxe els mercats de les ciutats més grans.[5][141][155] La majoria de la població local de l'entorn rural no entén ni el significat ni les connotacions d'«espècie en perill d'extinció», ni que la caça de lèmurs és il·legal o que els lèmurs només es poden trobar a Madagascar. [156] Molts malgaixos tenen tabús (fady) sobre menjar-se'ls o caçar-los, però això no n'impedeix la caça a moltes regions.[12] Tot i que la caça sempre ha sigut una amenaça per a les poblacions de lèmurs, recentment s'ha tornat molt més seriosa a causa del deteriorament de les condicions socioeconòmiques.[141] Les privacions econòmiques han fet que la població es mogui pel país a la cerca de feina, cosa que provoca la pèrdua o el deteriorament de les tradicions locals.[63][142][156] La sequera i la gana també poden reduir l'eficàcia dels fady que protegeixen els lèmurs.[63] Les espècies més grosses, com els sifaques i els lèmurs de collar, són l'objectiu habitual, però les espècies més petites també són caçades o atrapades per accident en trampes col·locades per a preses més grosses.[5][142] Les partides de caçadors amb experiència, organitzats i que utilitzen armes de foc, fones i sarbatanes poden matar entre vuit i vint lèmurs en un dia. Les partides de caça organitzades i els paranyers es poden veure tant a zones no protegides com a racons remots d'àrees protegides.[63] Els parcs nacionals i altres àrees protegides no estan prou protegides per les agències encarregades de vetllar pel compliment de la llei,[156] sovint hi ha massa pocs guàrdies per cobrir grans àrees i a vegades el terreny del parc és massa accidentat per poder controlar-lo amb regularitat.[157]
Alguns lèmurs, com el lèmur coronat i altres espècies que han sigut mantingudes amb èxit en captivitat, a vegades són capturats per la població de Madagascar com a animals de companyia exòtics.[47][134] Els lèmurs grisos també són utilitzats com a animals domèstics,[134] tot i que només poden sobreviure en aquesta situació durant uns dos mesos.[158] La captura d'exemplars vius per al comerç d'animals de companyia exòtics amb països més rics no se sol considerar una amenaça a causa de les estrictes regulacions que en controlen l'exportació.[134][142]
Conservació
[modifica]
Els lèmurs han atret molt l'atenció cap a Madagascar i les seves espècies en perill. Per això, aquests primats són espècies insígnia[N 8][63][134] de forma més notable ho fa el lèmur de cua anellada, que es considera una icona del país.[54] La presència de lèmurs als parcs nacionals atrau el turisme ecològic,[134] que ajuda especialment les comunitats rurals que viuen al voltant dels parcs nacionals perquè els ofereix oportunitats de treball i econòmiques, car la comunitat rep la meitat de les quotes del parc. En el cas del Parc Nacional de Ranomafana, les oportunitats de treball i els altres ingressos generats per la investigació a llarg termini poden rivalitzar amb les de l'ecoturisme.[160]
Des de l'any 1927 el govern de Madagascar ha declarat tots els lèmurs com a «protegits»[62] i ha establert àrees protegides que actualment es classifiquen en tres categories: Parcs Nacionals (Parcs Nationaux), Reserves Naturals Integrals (Réserves Naturelles Intégrales) i Reserves Especials (Réserves Spéciales). A principis del 2011 hi havia 21 parcs nacionals (19 de terrestres i 2 de marítims), 5 reserves naturals integrals i 21 reserves especials,[161] així com algunes petites reserves privades, com la Reserva de Berenty i la Reserva Privada de Sainte Luce, ambdues prop de Tôlanaro (anteriorment Fort-Dauphin).[134] Les àrees protegides, sense comptar les reserves privades, ocupen un 3% de la superfície de Madagascar i són administrades per l'organisme Madagascar National Parks, abans conegut com a Association Nationale pour la Gestion des Aires Protégées, així com altres organitzacions no governamentals com ara Conservation International, la Wildlife Conservation Society i el Fons Mundial per a la Natura.[134][144] La majoria d'espècies de lèmurs estan cobertes per aquesta xarxa d'àrees protegides i algunes d'elles poden trobar-se en diversos parcs o reserves.[144]
El Madagascar Fauna Group (MFG), una associació formada per prop de quaranta zoològics i organitzacions relacionades, és una altra de les entitats que vetllen per la conservació de la fauna malgaixa. Aquesta organització internacional no governamental col·labora en el manteniment del Parc Zoològic d'Ivoloina, ajuda a protegir la Reserva natural integral de Betampona i altres àrees protegides i promou la investigació de camp, per a la qual desenvolupa programes d'actuació, plans de conservació i programes educatius als zoos.[162] Un dels seus principals projectes suposà l'alliberament de lèmurs de collar blancs i negres captius, un projecte dissenyat per ajudar a reflotar la minvant població de la Reserva de Betampona.[162][163]

Els corredors ecològics són necessaris per unir aquestes àrees protegides de manera que les poblacions petites no quedin aïllades.[144] El setembre del 2003 a Durban (Sud-àfrica) l'expresident de Madagascar Marc Ravalomanana prometé triplicar la mida de les àrees protegides de l'illa en cinc anys.[141] Aquesta declaració es coneix com a «Visió de Durban».[134] L'any 1990 les selves pluvials d'Atsinanana, que inclouen sis parcs nacionals localitzats a la costa oriental de l'illa, i el juny del 2007 la Reserva natural integral de Tsingy de Bemaraha, que comprèn una part important de les selves tropicals de l'est de Madagascar, foren catalogades com a bé natural Patrimoni de la Humanitat de la UNESCO.[31][164]
La condonació de deute exterior pot ajudar Madagascar a protegir la seva biodiversitat.[151] Amb la crisi política del 2009, la tala il·legal ha proliferat i amenaça les selves tropicals del nord-est, fet que posa en perill els lèmurs que les habiten i el turisme ecològic del qual depenen les comunitats locals.[150]
Es mantenen algunes poblacions en captivitat a molts zoològics fora de Madagascar, tot i que la diversitat d'espècies és baixa. Els sifaques, per exemple, no sobreviuen gaire en captivitat, per la qual cosa poques instal·lacions tenen exemplars d'aquests animals.[12][59] La població en captivitat més nombrosa es troba al Duke Lemur Center, la missió del qual inclou la investigació no invasiva, la conservació i reproducció en captivitat i l'educació orientada cap al públic.[134]
Aspectes culturals
[modifica]En la cultura malgaixa, els lèmurs i els animals en general tenen ànima (ambiroa), que pot buscar venjança si es burlen d'ella mentre l'animal està viu o si el maten de manera cruel. Per això, els lèmurs, com molts altres elements de la vida quotidiana, han sigut font de tabús coneguts localment com a fady, que es basen en històries que giren al voltant de quatre principis bàsics: un poble o una regió poden creure que un cert tipus de lèmur pot ser l'avantpassat del clan, que l'esperit d'un lèmur pot buscar venjança, que l'animal pot ser un benefactor o que els lèmurs transmeten les seves qualitats, bones o dolentes, als bebès humans.[165] En general, els fady s'estenen més enllà d'allò que està prohibit i poden incloure esdeveniments que porten mala sort.[75]

Els tabús poden ajudar a protegir els lèmurs i els seus boscos en circumstàncies socioeconòmiques estables, però també poden conduir a la seva discriminació i persecució si es creu que un lèmur porta mala sort, per exemple, si camina per la ciutat.[63][165] D'altra banda, els fady no protegeixen tots els lèmurs d'igual manera; així, per exemple, tot i que caçar i menjar-se certes espècies pot ser tabú, d'altres no gaudeixen de la mateixa protecció i es converteixen, per tant, en objectiu dels caçadors.[165][12] Els tabús poden variar d'un poble a l'altre dins de la mateixa regió.[62] Si la gent es trasllada a un nou poble o regió, el seu fady pot no aplicar-se a les espècies de lèmur presents a la localitat, de manera que l'individu no vegi cap problema en el seu consum. Les restriccions dels tabús respecte a la carn de lèmur poden relaxar-se en temps de fam i sequera.[63]
L'ai-ai és considerat un animal no favorable arreu de Madagascar, tot i que les històries varien de poble en poble i de regió en regió.[75] Si la gent veu un ai-ai, pot matar-lo i penjar-ne el cadàver en un pal prop d'un camí fora de la població (i així ser altres els que puguin endur-se la mala sort) o cremar el seu poble i mudar-se.[62][48] Entre les supersticions relatives als ai-ais hi ha la creença que maten i es mengen els pollastres o fins i tot les persones, que maten humans mentre dormen,[63] que encarnen esperits d'ancestres[62] o que adverteixen de malalties, mort o mala sort en la família.[47][48] Al voltant del 1970, la població del districte de Marolambo (província de Toamasina) temia els ai-ais perquè creien que tenien poders sobrenaturals, per la qual cosa no permetien que ningú se'n burlés, els matés o se'ls mengés[165]
També hi ha un gran nombre de tabús relacionats amb l'indri i els sifaques. Sovint són protegits de la caça i el consum per la seva semblança amb els humans i els seus avantpassats, sobretot a causa de la seva gran talla i la seva postura erecta o ortògrada. La semblança és encara més notable en el cas de l'indri, que manca de la llarga cua que tenen la majoria de lèmurs actuals.[59][76] Conegut localment com a babakoto ('avantpassat de l'home'), a vegades és vist com a progenitor de la família o el clan. També hi ha històries d'un indri que ajudà un humà a baixar d'un arbre i sovint se'l veu com a benefactor.[165] Un altre fady es refereix a la creença que una dona tindrà nens lletjos si el seu marit mata un indri llanós o que, si una dona embarassada es menja un lèmur nan, el seu bebè n'adquirirà els ulls rodons i formosos.[165]
Els lèmurs també s'han fet populars en la cultura occidental recentment. La pel·lícula d'animació de l'any 2005 Madagascar fou vista per aproximadament 100 milions de persones als cinemes i 200-300 milions de persones en DVD arreu del món.[55] Abans d'aquesta pel·lícula, Zoboomafoo, una sèrie infantil de televisió de la PBS estatunidenca emesa entre el 1999 i el 2001,[166] ajudà a popularitzar els sifaques mostrant un sifaca de Coquerel del Duke Lemur Center.[167] Una sèrie de vint episodis titulada Lemur Street s'emeté el 2008 al canal Animal Planet; aquesta sèrie combinà el típic documental sobre animals amb la narració dramàtica per explicar la història de dos grups de lèmurs de cua anellada a la reserva privada de Berenty.[168][169][170][171]
Notes
[modifica]- ↑ No s'han trobat restes fòssils de cap exemplar jove de Mesopropithecus, Babakotia o Archaeoindris i se sap poca cosa sobre la dentadura caduca de Palaeopropithecus. Els models de desenvolupament es dedueixen dels dels seus parents més propers, els índrids.[72]
- ↑ En els índrids, una incisiva o canina inferiors no són substituïdes en la dentadura permanent. Existeixen interpretacions discrepants sobre aquesta producció de diferents fórmules dentals. Així, una fórmula dental alternativa per aquesta família és .[51]
- ↑ En primatologia, una organització de fissió-fusió és aquella en què la composició canvia cada dia, car els seus membres se separen durant períodes més o menys llargs. Per exemple, els individus dormen junts, però busquen aliment en petits grups que es desplacen en diferents direccions durant el dia.[92]
- ↑ Els grups multimascle, també anomenats multimascle/multifemella, són un tipus de organització social en què el grup està format per més d'un mascle adult, més d'una femella adulta i cries i joves de totes les edats. És el grup social més comú entre els primats i està format per entre 10 i 100 individus.[94]
- ↑ El salt i agafament vertical és el tipus de locomoció arbòria més estès entre els primats estrepsirrins, sobretot entre els índrids. En repòs, l'animal s'agafa a un suport vertical, com ara el lateral d'un arbre o una canya de bambú, i es desplaça d'una planta a l'altra saltant amb els membres inferiors i agafant-se a un altre suport vertical.[105]
- ↑ La criatura que descrigué Flacourt es considerà tradicionalment una espècie de Megaladapis. Tanmateix, estudis més recents apunten que probablement era una espècie de paleopropitec.[132]
- ↑ El raonament transitiu és la capacitat que té un individu d'inferir una relació entre dos o més elements que no s'han presentat junts. És una forma de raonament deductiu que s'ha suggerit com a mecanisme cognoscitiu pel qual els animals podrien aprendre les múltiples relacions dins de la jerarquia de domini del seu grup.
- ↑ Les espècies insígnia (flagship species) o totèmiques són espècies que s'elegeixen per representar una causa ambiental, com ara un ecosistema que necessita conservació, i que s'escullen per la seva vulnerabilitat, el seu atractiu o la seva peculiaritat per a generar suport i reconeixement del públic en general i així, amb el suport a aquestes espècies, aconseguir la conservació d'un ecosistema complet i de tots els animals que hi viuen.[159]
Referències
[modifica]- ↑ «The Paleobiology Database. Informació taxonòmica i de distribució sobre plantes i animals fòssils.» (en anglès).
- ↑ Von Linné, C. Systema Naturae (en anglès). 1. Estocolm: Impensis Direct. Laurentii Salvii, 1758, p. 29-30.
- ↑ Tattersall 1982, pàg. 43-44
- ↑ «Entrada «lèmur»». Diccionari.cat. [Consulta: 28 agost 2011].
- ↑ 5,0 5,1 5,2 5,3 5,4 5,5 5,6 5,7 5,8 Garbutt 2007, pàg. 85-86
- ↑ Lux, J «What are lemures?» (en anglès). Humanitas, 32, 1, 2008, pàg. 7-14. Arxivat de l'original el 2010-12-26 [Consulta: 28 agost 2011]. Arxivat 2010-06-06 a Wayback Machine.
- ↑ Flower & Lydekker 1891, p. 682
- ↑ Nield 2007, p. 41
- ↑ Blunt & Stearn 2002, p. 252
- ↑ 10,0 10,1 Kay, R. F.; Ross, C.; Williams, B. A «Anthropoid origins» (en anglès). Science, 275, 5.301, 1997, pàg. 797-804. DOI: 10.1126/science.275.5301.797. PMID: 9012340.
- ↑ 11,0 11,1 11,2 Gould & Sauther 2006, pàg. vii–xiii
- ↑ 12,00 12,01 12,02 12,03 12,04 12,05 12,06 12,07 12,08 12,09 12,10 12,11 12,12 12,13 12,14 12,15 12,16 12,17 12,18 12,19 12,20 12,21 12,22 12,23 12,24 12,25 12,26 12,27 12,28 12,29 12,30 12,31 12,32 12,33 12,34 12,35 12,36 12,37 12,38 12,39 12,40 Sussman 2003, pàg. 149-229
- ↑ 13,00 13,01 13,02 13,03 13,04 13,05 13,06 13,07 13,08 13,09 13,10 13,11 13,12 13,13 13,14 13,15 13,16 13,17 13,18 13,19 13,20 Ankel-Simons 2007, pàg. 392-514
- ↑ 14,00 14,01 14,02 14,03 14,04 14,05 14,06 14,07 14,08 14,09 14,10 14,11 14,12 14,13 14,14 14,15 14,16 Preston-Mafham 1991, pàg. 141-188
- ↑ 15,0 15,1 15,2 15,3 15,4 15,5 15,6 15,7 Tattersall 2006, pàg. 3-18
- ↑ 16,0 16,1 16,2 16,3 16,4 16,5 Yoder 2003, pàg. 1.242-1.247
- ↑ Yoder, A. D.; Yang, Z «Divergence dates for Malagasy lemurs estimated from multiple gene loci: geological and evolutionary context» (en anglès). Molecular Ecology, 13, 4, 2004, pàg. 757-773. DOI: 10.1046/j.1365-294X.2004.02106.x. PMID: 15012754.
- ↑ 18,0 18,1 18,2 Godinot, M «Lemuriform origins as viewed from the fossil record» (en anglès). Folia Primatologica, 77, 6, 2006, pàg. 446-464. DOI: 10.1159/000095391. PMID: 17053330.
- ↑ Flynn & Wyss 2003, pàg. 34-40
- ↑ 20,0 20,1 20,2 20,3 Mittermeier et al. 2006, pàg. 23-26
- ↑ Matthew, W. D «Climate and evolution» (en anglès). Annals of the New York Academy of Sciences, 24, 1915, pàg. 171-318. DOI: 10.1111/j.1749-6632.1914.tb55346.x.
- ↑ Garbutt 2007, pàg. 14-15
- ↑ Kline, G. «Study: Animals populated Madagascar by rafting there» (en anglès). Purdue Newsroom, 20-01-2010. [Consulta: 31 desembre 2010].
- ↑ 24,0 24,1 24,2 Horvath, J. E [et al]. «Development and application of a phylogenomic toolkit: Resolving the evolutionary history of Madagascar's lemurs» (en anglès). Genome Research, 18, 3, 2008, pàg. 490. DOI: 10.1101/gr.7265208. PMC: 2259113. PMID: 18245770.
- ↑ Krause 2003, pàg. 40-47
- ↑ 26,0 26,1 Ali, J.R.; Huber, M «Mammalian biodiversity on Madagascar controlled by ocean currents» (en anglès). Nature, 463, 7281, 2010, pàg. 653-656. DOI: 10.1038/nature08706. PMID: 20090678.
- ↑ 27,00 27,01 27,02 27,03 27,04 27,05 27,06 27,07 27,08 27,09 27,10 27,11 27,12 27,13 27,14 Sussman 2003, pàg. 107-148
- ↑ 28,0 28,1 28,2 28,3 28,4 28,5 28,6 Mittermeier et al. 2006, pàg. 89-182
- ↑ 29,0 29,1 29,2 29,3 Mittermeier et al. 2006, pàg. 37-51
- ↑ 30,00 30,01 30,02 30,03 30,04 30,05 30,06 30,07 30,08 30,09 Godfrey, Jungers & Schwartz 2006, pàg. 41-64
- ↑ 31,0 31,1 31,2 31,3 31,4 31,5 31,6 31,7 31,8 Yoder, A.D. «Lemurs: a quick guide» (en anglès). Current Biology, 17, 20, 2007, pàg. 866-868.
- ↑ 32,0 32,1 Dunham, A. E.; Rudolf, V. H. W «Evolution of sexual size monomorphism: the influence of passive mate guarding» (en anglès). Journal of Evolutionary Biology, 22, 7, 2009, pàg. 1376-1386. Arxivat de l'original el 2020-03-26. DOI: 10.1111/j.1420-9101.2009.01768.x. PMID: 19486235 [Consulta: 8 octubre 2011]. Arxivat 2020-03-26 a Wayback Machine.
- ↑ Preston-Mafham 1991, pàg. 10-21
- ↑ 34,0 34,1 34,2 Gommery, D.; Ramanivosoa, B.; Tombomiadana-Raveloson, S. [et al]. «A new species of giant subfossil lemur from the North-West of Madagascar (Palaeopropithecus kelyus, Primates)» (en anglès). Comptes Rendus Palevol, 3, 5, 2009, pàg. 471-480. DOI: 10.1016/j.crpv.2009.02.001.
- ↑ 35,0 35,1 35,2 Burney 2003, pàg. 47-51
- ↑ 36,00 36,01 36,02 36,03 36,04 36,05 36,06 36,07 36,08 36,09 36,10 36,11 36,12 36,13 36,14 36,15 36,16 36,17 36,18 36,19 36,20 36,21 36,22 36,23 36,24 36,25 36,26 36,27 36,28 Sussman 2003, pàg. 257-269
- ↑ 37,0 37,1 37,2 Godfrey & Jungers 2003, pàg. 1247-1252
- ↑ 38,0 38,1 38,2 Orlando, L. [et al]. «DNA from extinct giant lemurs links archaeolemurids to extant indriids» (en anglès). BMC Evolutionary Biology, 8, 121, 2008. DOI: 10.1186/1471-2148-8-121. PMC: 2386821. PMID: 18442367.
- ↑ 39,0 39,1 39,2 39,3 39,4 39,5 39,6 39,7 39,8 Rowe 1996, p. 27
- ↑ Groeneveld, L. F.; Weisrock, D. W.; Rasoloarison, R. M. [et al]. «Species delimitation in lemurs: multiple genetic loci reveal low levels of species diversity in the genus Cheirogaleus» (en anglès). BMC Evolutionary Biology, 9, 30, 2009. DOI: 10.1186/1471-2148-9-30. PMC: 2652444. PMID: 19193227.
- ↑ 41,0 41,1 41,2 Tattersall, I «Madagascar's lemurs: Cryptic diversity or taxonomic inflation?» (en anglès). Evolutionary Anthropology, 16, 2007, pàg. 12-23. DOI: 10.1002/evan.20126.
- ↑ 42,0 42,1 42,2 42,3 Mittermeier, R.; Ganzhorn, J.; Konstant, W. [et al]. «Lemur diversity in Madagascar» (en anglès). International Journal of Primatology, 29, 6, 2008, pàg. 1607-1656. Arxivat de l'original el 2018-08-27. DOI: 10.1007/s10764-008-9317-y [Consulta: 16 octubre 2011]. Arxivat 2018-08-27 a Wayback Machine.
- ↑ Karanth, P.; Rakotosamimanana, B.; Parsons, T. J. [et al]. «Ancient DNA from giant extinct lemurs verifies single origin of Malagasy primates» (en anglès). Proceedings of the National Academy of Sciences, 102, 14, 2005, pàg. 5090-5095. DOI: 10.1073/pnas.0408354102. PMC: 555979. PMID: 15784742.
- ↑ 44,0 44,1 Groves (2005)
- ↑ Cartmill 2010, pàg. 10-30
- ↑ 46,0 46,1 46,2 46,3 46,4 Sterling & McCreless 2006, pàg. 159-184
- ↑ 47,00 47,01 47,02 47,03 47,04 47,05 47,06 47,07 47,08 47,09 47,10 47,11 47,12 Ankel-Simons 2007, pàg. 48-161
- ↑ 48,0 48,1 48,2 Garbutt 2007, pàg. 205-207
- ↑ 49,0 49,1 Yoder, A.D.; Vilgalys, R.; Ruvolo, M «Molecular evolutionary dynamics of cytochrome b in strepsirrhine primates: The phylogenetic significance of third-position transversions» (en anglès). Molecular Biology and Evolution, 13, 10, 1996, pàg. 1339-1350. ISSN: 0737-4038. PMID: 8952078.
- ↑ Marivaux, L.; Welcomme, J.-L.; Antoine, P.-O.; Métais, G.; Baloch, I. M.; Benammi, M.; Chaimanee, Y.; Ducrocq, S.; Jaeger, J.-J «A fossil lemur from the Oligocene of Pakistan» (en anglès). Science, 294, 5542, 2001, pàg. 587-591. DOI: 10.1126/science.1065257. PMID: 11641497.
- ↑ 51,0 51,1 51,2 51,3 51,4 51,5 51,6 51,7 51,8 Ankel-Simons 2007, pàg. 224-283
- ↑ 52,0 52,1 Garbutt 2007, pàg. 115-136
- ↑ 53,0 53,1 Mittermeier et al. 2006, pàg. 209-323
- ↑ 54,0 54,1 54,2 Garbutt 2007, pàg. 137-175
- ↑ 55,0 55,1 Mittermeier et al. 2006, pàg. 85-88
- ↑ 56,0 56,1 56,2 Garbutt 2007, pàg. 176-204
- ↑ 57,0 57,1 Thalmann 2003, pàg. 1.340-1.342
- ↑ 58,0 58,1 58,2 58,3 Thalmann & Ganzhorn 2003, pàg. 1.336-1.340
- ↑ 59,0 59,1 59,2 59,3 59,4 59,5 59,6 59,7 Thalmann & Powzyk 2003, pàg. 1342-1345
- ↑ 60,0 60,1 60,2 Nowak 1999, pàg. 84-89
- ↑ 61,0 61,1 61,2 Richard 2003, pàg. 1.345-1.348
- ↑ 62,0 62,1 62,2 62,3 62,4 Sterling 2003, pàg. 1.348-1.351
- ↑ 63,00 63,01 63,02 63,03 63,04 63,05 63,06 63,07 63,08 63,09 63,10 63,11 63,12 Goodman, Ganzhorn & Rakotondravony 2003, pàg. 1159-1186
- ↑ 64,0 64,1 64,2 64,3 64,4 64,5 Ankel-Simons 2007, pàg. 284-391
- ↑ 65,0 65,1 65,2 65,3 Rowe 1996, p. 13
- ↑ Sellers, W. I. «Strepsirhine Haplorhine Split» (en anglès). Primate Adaptation and Evolution, 1999. [Consulta: 9 gener 2011].[Enllaç no actiu]
- ↑ 67,0 67,1 67,2 67,3 67,4 Overdorff & Johnson 2003, pàg. 1320-1324
- ↑ 68,0 68,1 Thalmann, U «Lemurs – ambassadors for Madagascar» (en anglès). Madagascar Conservation & Development, 1, 2006, pàg. 4-8. ISSN: 1662-2510.
- ↑ 69,0 69,1 Lamberton, C. «Dentition de lait de quelques lémuriens subfossiles malgaches» (en francès). Mammalia, 2, 1938, pàg. 57-80.
- ↑ Mittermeier et al. 1994, pàg. 33-48
- ↑ 71,0 71,1 Godfrey & Jungers 2002, pàg. 108-110
- ↑ Godfrey, Petto & Sutherland 2001, pàg. 113-157
- ↑ 73,00 73,01 73,02 73,03 73,04 73,05 73,06 73,07 73,08 73,09 Cuozzo & Yamashita 2006, pàg. 67-96
- ↑ 74,0 74,1 74,2 74,3 74,4 74,5 Powzyk & Mowry 2006, pàg. 353-368
- ↑ 75,0 75,1 75,2 Simons, E.L.; Meyers, D.M. «Folklore and beliefs about the Aye aye (Daubentonia madagascariensis)» (en anglès). Lemur News, 6, 2001, pàg. 11-16. Arxivat de l'original el 2011-07-23. ISSN: 0343-3528 [Consulta: 8 octubre 2013]. Arxivat 2011-07-23 a Wayback Machine.
- ↑ 76,0 76,1 Irwin 2006, pàg. 305-326
- ↑ Ankel-Simons 2007, pàg. 206-223
- ↑ 78,0 78,1 Ankel-Simons 2007, pàg. 162-205
- ↑ 79,0 79,1 79,2 79,3 79,4 79,5 79,6 79,7 Ankel-Simons 2007, pàg. 521-532
- ↑ 80,0 80,1 80,2 Bolwig, N. «Observations and thoughts on the evolution of facial mimic» (en anglès). Koedoe – African Protected Area Conservation and Science, 2, 1, 1959, pàg. 60-69. DOI: 10.4102/koedoe.v2i1.854.
- ↑ 81,0 81,1 Ross & Kay 2004, pàg. 3-41
- ↑ 82,0 82,1 82,2 82,3 Kirk & Kay 2004, pàg. 539-602
- ↑ 83,0 83,1 83,2 83,3 83,4 83,5 Jolly 2003, pàg. 1329-1331
- ↑ 84,0 84,1 84,2 84,3 84,4 84,5 84,6 84,7 84,8 Schmid & Stephenson 2003, pàg. 1198-1203
- ↑ 85,0 85,1 Nowak 1999, pàg. 65-71
- ↑ 86,0 86,1 86,2 Fietz 2003, pàg. 1307-1309
- ↑ 87,0 87,1 87,2 87,3 87,4 87,5 87,6 Birkinshaw & Colquhoun 2003, pàg. 1207-1220
- ↑ 88,00 88,01 88,02 88,03 88,04 88,05 88,06 88,07 88,08 88,09 Wright 2006, pàg. 385-402
- ↑ Nakajima, Y.; Shantha, T. R.; Bourne, G. H «Histochemical detection of l-gulonolactone: phenazine methosulfate oxidoreductase activity in several mammals with special reference to synthesis of vitamin C in primates» (en anglès). Histochemistry and Cell Biology, 18, 4, 1969, pàg. 293-301. DOI: 10.1007/BF00279880.
- ↑ Williams, C.V.; Campbell, J.L.; Glenn, K. M. «Comparison of serum iron, total iron binding capacity, ferritin, and percent transferrin saturation in nine species of apparently healthy captive lemurs» (en anglès). American Journal of Primatology, 68, 5, 2006, pàg. 1-13. DOI: 10.1002/ajp.20237. PMID: 16550526.
- ↑ Glenn, K.M.; Campbell, J.L.; Botstein, D.; Williams, C. V. «Retrospective evaluation of the incidence and severity of hemosiderosis in a large captive lemur population» (en anglès). American Journal of Primatology, 68, 4, 2006, pàg. 369-381. DOI: 10.1002/ajp.20231. PMID: 16534809.
- ↑ Sarmiento, F. O.; Vera, F.; Juncosa, J.. Diccionario de ecología: paisajes, conservación y desarrollo sustentable para Latinoamérica (en castellà). Editorial Abya Yala, 2000, p. 88. ISBN 9978046771.
- ↑ 93,0 93,1 Sussman 2003, pàg. 3-37
- ↑ Sussman 2003, p. 29
- ↑ Jolly, A. «Pair-bonding, female aggression and the evolution of lemur societies» (en anglès). Folia Primatologica, 69, 1998, pàg. 1-13. DOI: 10.1159/000052693.
- ↑ 96,0 96,1 96,2 96,3 Dunham, A.E. «Battle of the sexes: Cost asymmetry explains female dominance in lemurs» (en castellà). Animal Behavior, 76, 2008, pàg. 1435-1439. Arxivat de l'original el 2015-06-21. DOI: 10.1016/j.anbehav.2008.06.018 [Consulta: 8 abril 2014].
- ↑ 97,0 97,1 Young, A.L.; Richard, A.F.; Aiello, L.C. «Female dominance and maternal investment in strepsirhine primates» (en castellà). The American Naturalist, 135, 1990, pàg. 473-488. DOI: 10.1086/285057.
- ↑ Tan 2006, pàg. 369-382
- ↑ Digby, L.I.; Kahlenberg, S.M. «Female dominance in blue-eyed black lemurs» (en castellà). Primates, 43, 2002, pàg. 191-199. DOI: 10.1007/BF02629647. PMID: 12145400.
- ↑ 100,0 100,1 100,2 100,3 100,4 Jolly, A. «Lemur social behavior and primate intelligence» (en anglès). Science, 153, 3735, 1966, pàg. 501-506. DOI: 10.1126/science.153.3735.501.
- ↑ 101,0 101,1 Curtis 2006, pàg. 133-158
- ↑ Johnson 2006, pàg. 187-210
- ↑ 103,0 103,1 103,2 103,3 Fietz & Dausmann 2006, pàg. 97-110
- ↑ Garbutt 2007, pàg. 86-114
- ↑ Sussman 2003, p. 78
- ↑ Godfrey, L.R.; Jungers, W.L. «The extinct sloth lemurs of Madagascar» (en anglès). Evolutionary Anthropology, 12, 2003, pàg. 252-263. DOI: 10.1002/evan.10123.
- ↑ Nowak 1999, pàg. 71-81
- ↑ Mittermeier et al. 2006, pàg. 324-403
- ↑ Sauther, M. L «Antipredator behavior in troops of free-ranging Lemur catta at Beza Mahafaly special reserve, Madagascar» (en anglès). International Journal of Primatology, 10, 6, 1989, pàg. 595-606. DOI: 10.1007/BF02739366.
- ↑ Jolly 1966, p. 135
- ↑ Grieser, B «Infant development and parental care in two species of sifakas» (en anglès). Primates, 33, 3, 1992, pàg. 305-314. DOI: 10.1007/BF02381192.
- ↑ 112,0 112,1 112,2 Vasey 2003, pàg. 1332-1336
- ↑ Braune, P.; Schmidt, S.; Zimmermann, E «Acoustic divergence in the communication of cryptic species of nocturnal primates (Microcebus ssp.)» (en anglès). BMC Biology, 6, 19, 2008. DOI: 10.1186/1741-7007-6-19. PMC: 2390514. PMID: 18462484.
- ↑ 114,0 114,1 G. R., Hosey; Thompson, R. J «Grooming and touching behaviour in captive ring-tailed lemurs (Lemur catta L.)» (en anglès). Primates, 26, 1, 1985, pàg. 95-98. DOI: 10.1007/BF02389051.
- ↑ Barton, R. A «Allogrooming as mutualism in diurnal lemurs» (en anglès). Primates, 28, 4, 1987, pàg. 539-542. DOI: 10.1007/BF02380868.
- ↑ 116,0 116,1 116,2 116,3 116,4 116,5 Goodman 2003, pàg. 1221-1228
- ↑ Eberle, M.; Kappele, P.M. «Mutualism, reciprocity, or kin selection? Cooperative rescue of a conspecific from a boa in a nocturnal solitary forager the Gray Mouse Lemur» (en anglès). American Journal of Primatology, 70, 4, 2008, pàg. 410-414. Arxivat de l'original el 2011-07-19. DOI: 10.1002/ajp.20496. PMID: 17972271 [Consulta: 7 octubre 2015]. Arxivat 2011-07-19 a Wayback Machine.
- ↑ Crompton & Sellers 2007, pàg. 127-145
- ↑ 119,0 119,1 Kappeler & Rasoloarison 2003, pàg. 1.310-1.315
- ↑ 120,0 120,1 Mutschler & Tan 2003, pàg. 1324-1329
- ↑ 121,0 121,1 Patel, E. R «Non-maternal infant care in wild Silky Sifakas (Propithecus candidus)» (en anglès). Lemur News, 12, 2007, pàg. 39-42. Arxivat de l'original el 2011-07-23. ISSN: 1608-1439 [Consulta: 10 octubre 2015]. Arxivat 2011-07-23 a Wayback Machine.
- ↑ J., Fietz; Dausmanna, K. H «Costs and Potential Benefits of Parental Care in the Nocturnal Fat-Tailed Dwarf Lemur (Cheirogaleus medius)» (en anglès). Folia Primatologica, 74, 5-6, 2003, pàg. 246-258. DOI: 10.1159/000073312. PMID: 14605471.
- ↑ Vasey, N «The breeding system of wild red ruffed lemurs (Varecia rubra): a preliminary report» (en anglès). Primates, 48, 1, 2007, pàg. 41-54. DOI: 10.1007/s10329-006-0010-5. PMID: 17024514.
- ↑ Ehrlich, A.; Fobes, J.; King, J «Prosimian learning capacities» (en anglès). Journal of Human Evolution, 5, 6, 1976, pàg. 599-617. DOI: 10.1016/0047-2484(76)90005-1.
- ↑ Fichtel & Kappeler 2010, pàg. 413
- ↑ Penn State. «Piecing together an extinct lemur, large as a big baboon» (en anglès). ScienceDaily, 29-07-2008. [Consulta: 30 desembre 2010].
- ↑ 127,0 127,1 127,2 127,3 127,4 Jolly & Sussman 2006, pàg. 19-40
- ↑ Bons, N.; Rieger, F.; Prudhomme, D.; Fisher, A.; Krause, K. H «Microcebus murinus: a useful primate model for human cerebral aging and Alzheimer's disease?» (en anglès). Genes, Brain and Behavior, 5, 2, 2006, pàg. 120-130. DOI: 10.1111/j.1601-183X.2005.00149.x. PMID: 16507003.
- ↑ Marchal, S.; Givalois, L.; Verdier, J. M.; Mestre-Francés, N «Distribution of lithostathine in the mouse lemur brain with aging and Alzheimer's-like pathology» (en anglès). Neurobiology of Aging, 2011. DOI: 10.1016/j.neurobiolaging.2011.01.002. PMID: 21371784.
- ↑ Gifford, R. J. et al «A transitional endogenous lentivirus from the genome of a basal primate and implications for lentivirus evolution» (en anglès). Proceedings of the National Academy of Sciences, 105, 51, 2008, pàg. 20.362-7. DOI: 10.1073/pnas.0807873105. PMC: 2603253. PMID: 19075221.
- ↑ Mittermeier et al. 2006, p. 29
- ↑ Simons 1997, pàg. 142-166
- ↑ 133,0 133,1 133,2 Mittermeier et al. 2006, pàg. 27-36
- ↑ 134,00 134,01 134,02 134,03 134,04 134,05 134,06 134,07 134,08 134,09 134,10 134,11 134,12 Mittermeier et al. 2006, pàg. 52-84
- ↑ Macedonia, J. M «The vocal repertoire of the Ring-tailed Lemur (Lemur catta)» (en anglès). Folia Primatologica, 61, 1993, pàg. 186-217. DOI: 10.1159/000156749.
- ↑ Tilden, C. D «A study of locomotor behavior in a captive colony of red-bellied lemurs (Eulemur rubriventer)» (en anglès). American Journal of Primatology, 22, 2, 1990, pàg. 87-100. DOI: 10.1002/ajp.1350220203.
- ↑ Wunderlich, R. E. «Kinematics of bipedalism in Propithecus verreauxi» (en anglès). Journal of Zoology, 272, 2, 2007, pàg. 165-175. DOI: 10.1111/j.1469-7998.2006.00253.x.
- ↑ MacLean, E. L. «Social complexity predicts transitive reasoning in prosimian primates» (en anglès). Animal Behaviour, 76, 2, 2008, pàg. 479-486. DOI: 10.1016/j.anbehav.2008.01.025. PMC: 2598410. PMID: 19649139.
- ↑ Merritt, D. «A comparative analysis of serial ordering in Ring-Tailed Lemurs (Lemur catta)» (en anglès). Journal of Comparative Psychology, 121, 4, 2007, pàg. 363-371. DOI: 10.1037/0735-7036.121.4.363. PMC: 2953466. PMID: 18085919.
- ↑ Santos, Laurie R. «How prosimian primates represent tools: Experiments with two lemur species (Eulemur fulvus and Lemur catta)» (en anglès). Journal of Comparative Psychology, 119, 4, 2005, pàg. 394-403. DOI: 10.1037/0735-7036.119.4.394. PMID: 16366773.
- ↑ 141,0 141,1 141,2 141,3 141,4 141,5 Mittermeier et al. 2006, pàg. 15-17
- ↑ 142,0 142,1 142,2 142,3 142,4 Harcourt 1990, pàg. 7-13
- ↑ «UNEP-WCMC Species Database: CITES-Listed Species» (en anglès). UNEP-WCMC. [Consulta: 13 març 2011].[Enllaç no actiu]
- ↑ 144,0 144,1 144,2 144,3 144,4 144,5 144,6 Mittermeier, Konstant & Rylands 2003, pàg. 1538-1543
- ↑ Schwitzer, C.; Mittermeier, R. A.; Rylands, A. B.; Taylor, L. A.; Chiozza, F.; Williamson, E. A.; Wallis, J.; Clark, F. E (editors). «Primates in Peril: The World's 25 Most Endangered Primates 2012-2014» (en anglès) p. 1-91. IUCN/SSC Primate Specialist Group (PSG), Societat Primatològica Internacional (IPS), Conservation International (CI) i Bristol Zoological Society (BZS), 2012.
- ↑ Black, Richard «Lemurs sliding towards extinction» (en anglès). BBC News, 13-07-2012 [Consulta: 18 agost 2013].
- ↑ «91% of Madagascar’s lemurs threatened with extinction» (en anglès). Mongabay, 26-09-2012 [Consulta: 18 agost 2013].
- ↑ Samuel, H. «Furry lemurs 'could be wiped out within 20 years'» (en anglès). The Daily Telegraph, 19-08-2012 [Consulta: 18 agost 2013].
- ↑ Mintz, Z. «Lemurs Face Extinction In 20 Years, Risk Of Losing Species For 'First Time In Two Centuries'» (en anglès). International Business Times, 21-08-2013 [Consulta: 18 agost 2013].
- ↑ 150,0 150,1 150,2 «Country brief: Madagascar» (en anglès). eStandardsForum, 01-12-2009. Arxivat de l'original el 26 de juliol 2011. [Consulta: 14 gener 2011].
- ↑ 151,0 151,1 151,2 World Wildlife Fund. «Monumental debt-for-nature swap provides $20 million to protect biodiversity in Madagascar» (en anglès). ScienceDaily, 14-06-2008. [Consulta: 30 desembre 2010].
- ↑ Ganzhorn, Goodman & Dehgan 2003, pàg. 1.228-1.234
- ↑ Junge & Sauther 2006, pàg. 423-440
- ↑ Ratsimbazafy 2006, pàg. 403-422
- ↑ Butler, Rhett. «Madagascar's political chaos threatens conservation gains» (en anglès). Yale Environment 360. Yale School of Forestry & Environmental Studies, 04-01-2010. Arxivat de l'original el 26 de desembre 2010. [Consulta: 14 gener 2011].
- ↑ 156,0 156,1 156,2 Simons 1997, pàg. 142-166
- ↑ Schuurman, Derek «Illegal logging in Madagascar» (en anglès). Traffic Bulletin, 22, 2, 2009, pàg. 49.
- ↑ Rowe 1996, p. 46
- ↑ Sarmiento, F. O.; Vera, F. y Juncosa, J. Diccionario de ecología: paisajes, conservación y desarrollo sustentable para Latinoamérica (en castellà). Editorial Abya Yala, 2000, p. 88. ISBN 9978046771.
- ↑ Wright & Andriamihaja 2003, pàg. 1.485-1.488
- ↑ «Madagascar National Parks. Une structure, une mission» (en francès). Madagascar National Parks. Arxivat de l'original el 24 de maig 2011. [Consulta: 28 abril 2011].
- ↑ 162,0 162,1 Sargent & Anderson 2003, pàg. 1.543-1.545
- ↑ Britt et al. 2003, pàg. 1.545-1.551
- ↑ «Madagascar» (en anglès). World Heritage List. Unesco. [Consulta: 22 març 2011].
- ↑ 165,0 165,1 165,2 165,3 165,4 165,5 Ruud 1970, pàg. 97-101
- ↑ Barnes, D «Fun in the sun with lemurs» (en anglès). The Washington Times, 21-10-2001.
- ↑ Jacobson, Louis «Looking for lemurs» (en anglès). USA Today, 30-03-2004 [Consulta: 19 març 2011].
- ↑ Huff, R. «Tonight» (en anglès). New York Daily News, 08-02-2008, p. 122.
- ↑ «Highlights» (en anglès). The Washington Post, 08-02-2008, p. C04.
- ↑ «Madagascar; country's unknown creatures on DSTV» (en anglès). Africa News, 28-12-2007.
- ↑ Walmark, H «Critic's choice: Lemur Street» (en anglès). The Globe and Mail, 08-02-2008, p. R37.
Bibliografia
[modifica]- Ankel-Simons, F. Primate Anatomy (en anglès). 3a ed.. Academic Press, 2007. ISBN 0-12-372576-3.
- Blunt, W.; Stearn, W. T. Linnaeus: the compleat naturalist (en anglès). Princeton University Press, 2002. ISBN 978-0691096360.
- Flower, W. H.; Lydekker, R. An introduction to the study of mammals living and extinct (en anglès). Londres: A. and C. Black, 1891. ISBN 978-1110768578.
- Garbutt, N. Mammals of Madagascar: A Complete Guide (en anglès). A. and C. Black Publishers, 2007. ISBN 978-0-300-12550-4.
- Goodman, S. M.; Benstead, J. P.. The Natural History of Madagascar (en anglès). University of Chicago Press, 2003. ISBN 0-226-30306-3.
- Capítol 2 – «Geology and Soils»
- Flynn, J. J.; Wyss, A.R. Mesozoic Terrestrial Vertebrate Faunas: The Early History of Madagascar's Vertebrate Diversity, 2003, p. 34–40.
- Krause, D. W. Late Cretaceous Vertebrates of Madagascar: A Window into Gondwanan Biogeography at the End of the Age of Dinosaurs, 2003, p. 40–47.
- Burney, D. A. Madagascar's Prehistoric Ecosystems, 2003, p. 47–51.
- Capítol 13 – «Mammals»
- Goodman, S. M; Ganzhorn, J. U.; Rakotondravony, D. Introduction to the Mammals, 2003, p. 1159–1186.
- Schmid, J.; Stephenson, P. J. Physiological Adaptations of Malagasy Mammals: Lemurs and Tenrecs Compared, 2003, p. 1198–1203.
- Birkinshaw, C.R.; Colquhoun, I. C. Lemur Food Plants, 2003, p. 1207–1220.
- Goodman, S. M. Predation on Lemurs, 2003, p. 1221–1228.
- Ganzhorn, J. U.; Goodman, S. M.; Dehgan, A. Effects of Forest Fragmentation on Small Mammals and Lemurs, 2003, p. 1228–1234.
- Yoder, A. D.. Phylogeny of the Lemurs, 2003, p. 1242–1247.
- Godfrey, L. R.; Jungers, W. L.. Subfossil Lemurs, 2003, p. 1247–1252.
- Fietz, J. Primates: Cheirogaleus, Dwarf Lemurs or Fat-tailed Lemurs, 2003, p. 1307–1309.
- Kappeler, P. M.; Rasoloarison, R. M.. Microcebus, Mouse Lemurs, Tsidy, 2003, p. 1310–1315.
- Overdorff, D. J.; Johnson, S. Eulemur, True Lemurs, 2003, p. 1320–1324.
- Mutschler, T.; Tan, C. L.. Hapalemur, Bamboo or Gentle Lemur, 2003, p. 1324–1329.
- Jolly, A. Lemur catta, Ring-tailed Lemur, 2003, p. 1329–1331.
- Vasey, N. Varecia, Ruffed Lemurs, 2003, p. 1332–1336.
- Thalmann, U.; Ganzhorn, J. U.. Lepilemur, Sportive Lemur, 2003, p. 1336–1340.
- Thalmann, U. Avahi, Woolly Lemurs, 2003, p. 1340–1342.
- Thalmann, U.; Powzyk, J. Indri indri, Indri, 2003, p. 1342–1345.
- Richard, A. Propithecus, Sifakas, 2003, p. 1345–1348.
- Sterling, E. Daubentonia madagascariensis, Aye-aye, 2003, p. 1348–1351.
- Capítol 14 – «Conservation»
- Wright, P. C.; Andriamihaja, B. The conservation value of long-term research: a case study from the Parc National de Ranomafana, 2003, p. 1485–1488.
- Mittermeier, R. A.; Konstant, W.R.; Rylands, A.B.. Lemur Conservation, 2003, p. 1538–1543.
- Sargent, E. L.; Anderson, D. The Madagascar Fauna Group, 2003, p. 1543–1545.
- Britt, A.; Iambana, B. R.; Welch, C. R.; Katz, A. S.. Restocking of Varecia variegata variegata in the Réserve Naturelle Intégrale de Betampona, 2003, p. 1545–1551.
- Goodman, S. M. (ed.); Patterson, B. D. (ed.). Natural Change and Human Impact in Madagascar (en anglès). Smithsonian Institution Press, 1997. ISBN 978-1560986829.
- Simons, E. L.. Capítol 6: Lemurs: Old and New, 1997, p. 142–166.
- Gould, L. (ed.); Sauther, M. L. (ed.). Lemurs: Ecology and Adaptation (en anglès). Springer, 2006. ISBN 978-0387-34585-7.
- Gould, L.; Sauther, M. L.. Lemurs: Ecology and Adaptation, 2006, p. vii–xiii.
- Tattersall, I. Lemurs: Ecology and Adaptation, 2006, p. 3–18.
- Jolly, A.; Sussman, R. W.. Capítol 2: Notes on the History of Ecological Studies of Malagasy Lemurs, 2006, p. 19–40.
- Godfrey, L. R.; Jungers, W.L.; Schwartz, G. T.. Capítol 3: Ecology and Extinction of Madagascar's Subfossil Lemurs, 2006, p. 41–64.
- Cuozzo, F. P.; Yamashita, N. Capítol 4: Impact of Ecology on the Teeth of Extant Lemurs: A Review of Dental Adaptations, Function, and Life History, 2006, p. 67–96.
- Fietz, J.; Dausmann, K.H.. Capítol 5: Big Is Beautiful: Fat Storage and Hibernation as a Strategy to Cope with Marked Seasonality in the Fat-Tailed Dwarf Lemur (Cheirogaleus medius), 2006, p. 97–110.
- Curtis, D. J.. Capítol 7: Cathemerality in Lemurs, 2006, p. 133–158.
- Sterling, E. J.; McCreless, E.E.. Lemurs: Ecology and Adaptation, 2006, p. 159–184.
- Johnson, S. E.. Capítol 9: Evolutionary Divergence in the Brown Lemur Species Complex, 2006, p. 187–210.
- Irwin, M. T.. Capítol 14: Ecologically Enigmatic Lemurs: The Sifakas of the Eastern Forests (Propithecus candidus, P. diadema, P. edwardsi, P. perrieri, and P. tattersalli), 2006, p. 305–326.
- Powzyk, J. A.; Mowry, C. B.. Capítol 16: The Feeding Ecology and Related Adaptations of Indri indri, 2006, p. 353–368.
- Tan, C. L.. Capítol 17: Behavior and Ecology of Gentle Lemurs (Genus Hapalemur), 2006, p. 369–382.
- Wright, P. C.. Capítol 18: Considering Climate Change Effects in Lemur Ecology, 2006, p. 385–402.
- Ratsimbazafy, J. Capítol 19: Diet Composition, Foraging, and Feeding Behavior in Relation to Habitat Disturbance: Implications for the Adaptability of Ruffed Lemurs (Varecia variegata editorium) in Manombo Forest, Madagascar, 2006, p. 403–422.
- Junge, R. E.; Sauther, M. Capítul 20: Overview on the Health and Disease Ecology of Wild Lemurs: Conservation Implications, 2006, p. 423–440.
- Groves, Colin. Wilson, D. E.; Reeder, D. M. (editors). Mammal Species of the World (en anglès). 3a edició. Johns Hopkins University Press, 2005. ISBN 0-801-88221-4. (anglès)
- Gursky, K.A.I. (ed.); Nekaris, S.L. (ed.). Primate Anti-Predator Strategies (en anglès). Springer, 2009. ISBN 978-1441941909.
- Crompton, R. H.; Sellers, W. I.. A consideration of leaping locomotion as a means of predator avoidance in prosimian primates, 2007, p. 127–145.
- Harcourt, C. Thornback, J. (ed.). Lemurs of Madagascar and the Comoros: The IUCN Red Data Book (en anglès). Unió Internacional per a la Conservació de la Natura, 1990. ISBN 978-2880329570.
- Hartwig, W. C. (ed.). The primate fossil record (en anglès). Cambridge University Press, 2002. ISBN 978-0521663151.
- Godfrey, L. R.; Jungers, W. L.. Capítol 7: Quaternary fossil lemurs (en anglès), 2002, p. 97–121.
- Jolly, A. Lemur Behavior (en anglès). Chicago: University of Chicago Press, 1966. ISBN 978-0226405520.
- Mittermeier, R. A.; Tattersall, I.; Konstant, W. R. [et al.].. Lemurs of Madagascar (en anglès). Il·lustrat per S.D. Nash. 1a ed.. Conservation International, 1994. ISBN 1-881173-08-9.
- Mittermeier, R. A.; Konstant, W. R.; Hawkins, F. [et al.].. Lemurs of Madagascar (en anglès). Il·lustrat per S.D. Nash. 2a edició. Conservation International, 2006. ISBN 1-881173-88-7.
- Nield, T. Supercontinent: Ten Billion Years in the Life of Our Planet (en anglès). Harvard University Press, 2007. ISBN 978-0674026599.
- Nowak, R. M.. Walker's Mammals of the World (en anglès). 6a ed.. Johns Hopkins University Press, 1999. ISBN 0801857899.
- Platt, M. (ed.); Ghazanfar, A. (ed.). Primate Neuroethology (en anglès). Oxford University Press, 2010. ISBN 978-0195-32659-8.
- Cartmill, M. Primate neuroethology, 2010, p. 10–30. ISBN 9780195326598.
- Plavcan, J. M.; Kay, R.; Jungers, W. L.; van Schaik, C. (editors). Reconstructing behavior in the primate fossil record (en anglès). Springer, 2001. ISBN 978-0306466045.
- Godfrey, L. R.; Petto, A. J.; Sutherland, M. R.. Capítol 4: Dental ontogeny and life history strategies: The case of the giant extinct indroids of Madagascar, 2001, p. 113–157.
- Preston-Mafham, K. Madagascar: A Natural History (en anglès). Facts on File, 1991. ISBN 978-0816024032.
- Ross, C. F. (ed.); Kay, R. F. (ed.). Anthropoid Origins: New Visions (en anglès). 2a ed.. Springer, 2004. ISBN 978-0306481208.
- Ross, C. F.; Kay, R. F.. Capítol 1: Evolving Perspectives on Anthropoidea, 2004, p. 3–41.
- Kirk, C. E.; Kay, R. F.. Capítol 22: The Evolution of High Visual Acuity in the Anthropoidea, 2004, p. 539–602.
- Rowe, N. The Pictorial Guide to the Living Primates (en anglès). Pogonias Press, 1996. ISBN 978-0964882515.
- Ruud, J. Taboo: A Study of Malagasy Customs and Beliefs (en anglès). 2a ed.. Oslo University Press, 1970.
- Sussman, R. W.. Primate Ecology and Social Structure (en anglès). Pearson Custom Publishing, 2003. ISBN 978-0-536-74363-3.
- Tattersall, I. The Primates of Madagascar (en anglès). Columbia University Press, 1982. ISBN 0-231-04704-5.
Vegeu també
[modifica]Enllaços externs
[modifica]- Lemurs Arxivat 2018-08-31 a Wayback Machine. a BBC Nature (anglès)
- Lemurs of Madagascar (anglès)