Ekspresija gena
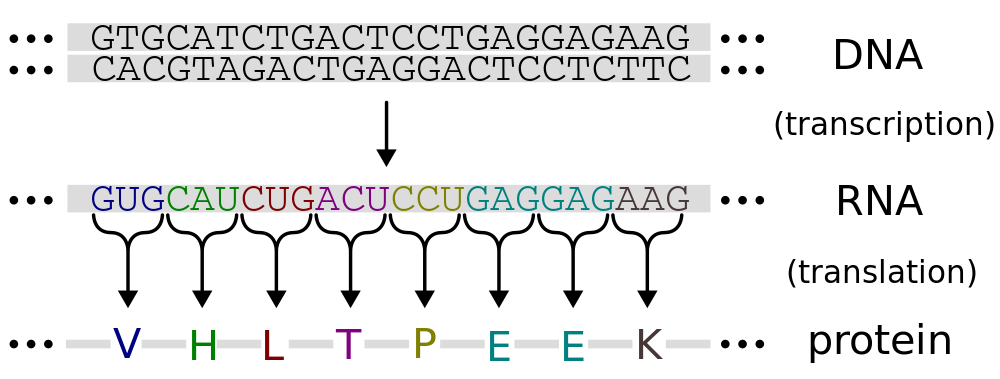
Ekspresija gena je proces kojim se informacija iz gena prepisuje i prevodi u funkcionalni genski produkt. Ovi proizvodi su obično proteini, ali i ne-proteinski kodirajući geni, kao što su tRNK ili snRNK (mala nuklearna RNK), proizvodi su funkcionalne RNK. Proces ekspresije gena je svojstven svim poznatim oblicima života - eukariota (uključujući i višećelijske organizme), prokariota (bakterija i archaea), a koje koriste virus - za generiranje makromolskih sistema za održavanje životnih struktura i funkcija.[1][2]
Nekoliko koraka u procesu genske ekspresije mogu biti modulirani, uključujući i transkripciju, preradu RNK, translaciju i post-modifikaciju proteina.
Regulacija ekspresije gena daje ćeliji mogućnost kontrole nad strukturom i funkcijom, a to je osnova za ćelijsku diferencijaciju, morfogenezu i svestranu prilagodljivost bilo kog organizma.
Genska regulacije može biti i osnov evolucijskih promjene, jer kontrola nad vremenom , mjestom i količinom ekspresije gena može imati dubok utjecaj na funkcije gena u ćeliji ili u višećelijskim organizmima.
U genetici, ekspresija gena je najfundamentalnijia razina na kojoj genotip utiče na formiranje fenotipa. Genetički kod iz molekula DNK interpretira se putem genske ekspresije, čije osobenosti, u interakciji sa epigenetičkom i spoljnom sredinom, formiraju individualni fenotip. Takvi fenotipovi su obično izraz sinteze proteina koji kontroliraju građu i funkciju organizma, odnosno djeluju kao njihovi regulatori – enzimi, koji kataliziraju specifične metaboličke puteve karakterizacije organizma.[3][4][5][6]
Mehanizmi
[uredi | uredi izvor]Prerada RNK, procesuiranje (splicing)
[uredi | uredi izvor]
Prerada RNK , eng. splicing = srastanje (egzona), je najznačajnija post-translacijska modifikacija primarnog transkripta i zrenja iRNK.
Dok transkripcija kodirajućih gena za sintezu prokariotskih proteina ide preko iRNK, transkripcija eukariotskih gena daje primarni transkript RNK (pre-iRNK), koji se mora proći procesuiranje u nizu modifikacija da postane zrela iRNK.
To uključuje 5' „kaptažu“, koja je uređena enzimskim reakcijama koje dodaju 7-metilguanozine (m7G) na 5' kraj pre-iRNK i na taj način štite RNK od degradacije putem egzonukleaza. Tako je 7G kaptažni faktor tada vezan u heterodimer (CBC20/CBC80), koji pomaže iRNK ulaz u citoplazmu i štite RNK od dekaptaže
Druga modifikacija je 3 otkrivanje i poliadenilacija. Ako je poliadenilacijski signal (5'AAUAAA-3') prisutan u pre-iRNK signal, obično se javljaju između sekvenci proteina i terminatora. Pre-iRNK se prvo rascijepi, a zatim se niz od približno 200 adenina (A) dodaje da formiraju poli (A) rep, koji RNK štiti od degradacije. Poli (A) rep je vezan više za proteine (PABP) neophodne za eksport iRNK i ponovno pokretanje translacije.
Većina eukariotskih pre-iRNK sastoji od naizmjeničnih segmenata zvanih egzoni i introni. U procesu prerade, RNK-protein katalizirajući kompleks poznat kao splajsosom, koji katalizira dvije trans-esterifikacijske reakcije, koje uklanjaju intron i oslobađaju ga u obliku omčaste (laso) strukture, a zatim uvezuju susjedne egzone. U određenim slučajevima, neki introni ili egzoni mogu biti uklonjeni ili zadržani u zrelim iRNK. Ova takozvana alternativna prerada (splicing) stvara niz različitih transkripata porijeklom iz jednog gena, jer te transkripte potencijalno može prevesti u različite proteine, čime prerada proširuje složenost ekspresije eukariotskih gena.
Regulacija ekspresije gena
[uredi | uredi izvor]Hromosom E. coli se sastoji od kružne molekule DNK (4,7 x 106 bp) koja kodira nekoliko hiljada različitih proteina, od kojih se samo neki sintetiziraju u određenom momentu. Neki proteini bakterijske ćelije su konstitutivni, tj. sintetiziraju se kontinuirano, nezavisno od trenutnih uslova, dok su drugi inducibilni i njihova sinteza je regulisana u skladu s intracelularnom koncentracijom pojedinih metabolita. Bakterije obično žive u vrlo promjenljivim uslovima, tako da enzimi koji su ćeliji potrebni u jednom trenutku ubrzo mogu postati suvišni, a iznenada se može pojaviti potreba za nekim drugim enzimima. Genom bakterija je organizovan tako da omogućuje veliku fleksibilnost u pogledu ekspresije pojedinih gena. Bakterija može da odgovori na promjene uslova veoma brzo, sintezom enzima koji su joj u datom trenutku neophodni, i prestankom sinteze onih za koje trenutno nema supstrata. Skoro kompletna regulacija ekspresije gena kod bakterija se odvija na nivou transkripcije. Geni bakterija su grupisani tako da se jedan do drugog nalaze oni koji kodiraju funkcionalno povezane enzime, najčešće enzime jednog biohemijskog puta. Cijeli set takvih gena sadrži samo jedan promotor i transkribuje se kao jedna transkripciona jedinica, a iRNK je policistronska. Set gena koji predstavlja jednu ekspresionu jedinicu, tj. uključuje strukturne gene i elemente koji kontrolišu njihovu ekspresiju, nazvan je operonom. Takva organizacija genoma omogućuje efikasnu i koordinisanu regulaciju seta gena, interakcijom između regulatornog proteina i zajedničkog promotora. Indukcija sinteze pojedinih enzima je veoma brza (obično traje samo nekoliko minuta) zahvaljujući i tome što su procesi transkripcije i translacije spregnuti. Translacija započinje na 5'-kraju iRNK čija je sinteza još u toku. Bakterijske iRNK su kratkoživeće, degradiraju već 1-3 minute poslije transkripcije čime se izbjegava sinteza onih proteina koji ćeliji više nisu potrebni. Osnovna saznanja o mehanizmima putem kojih se kontroliše aktivnost pojedinih gena u bakterijskim ćelijama pružila su ispitivanja regulacije sinteze inducibilnih enzima.



Regulacija transkripcije laktoznog operona - negativna kontrola
[uredi | uredi izvor]Prvi model regulacije ekspresije gena kod bakterija predložili su Jacob i Monod 1961. godine, na osnovu izučavanja sinteze enzima koji su uključeni u razgradnju laktoze kod bakterije E. coli. U bakterijskom genomu se nalaze tri strukturna gena (lacZ, lacY i lacA ) koji kodiraju glavne enzime za razgradnju laktoze: 0-galaktozidazu (katalizuje razgradnju laktoze do glukoze i gataktoze), P-galaktozid permeazu (omogućava transport laktoze kroz plazma membranu) i P-tiogalaktozid acetil-transferazu (fiziološka uloga joj nije utvrđena). Ova tri gena čine jednu transkripcionu jedinicu, tzv. laktozni operon (lac operon) čiji se promotor (lacP) nalazi uzvodno od gena lacZ. Uzvodno od promotora smješten je regulatorni gen (lac-1) koji predstavlja zasebnu transkripcionu jedinicu (ima svoj promotor) i transkribuje se u monocistronsku iRNK. Ova iRNK kodira protein koji ima ulogu represora transkripcije lac operona.
U toku ispitivanja mehanizma regulacije transkripcije lac operona, izolovan je i prečišćen laktozni represor (lac++ represor) – protein koji se vezuje za niz nukleotida nazvan operator. Ovaj niz sadrži 21 bp i prisutan je u genomu E. coli u samo jednoj kopiji, a lociran je unutar promotorskog regiona lacP. Laktozni represor je tetramer koji ima visok afinitet prema operatoru (lacO). Kada se represor veže za operator, on blokira pristup RNK polimerazi i sprečava transkripciju strukturnih gena vezanih za taj promotor. S druge strane, represor ima i vezivno mjesto za šećer alolaktozu (izomer D-laktoze koji je u ćeliji prisutan onda kada je prisutna i laktoza). Kada koncentracija alolaktoze u bakterijskoj ćeliji dostigne određeni nivo, ona se vezuje za represor, izazivajući njegovu alosteričnu promjenu, usljed koje slabi njegov afinitet prema operatoru, tako da represor disocira sa operatora. Sada je transkripcija gena ponovo omogućena. Ova pojava se naziva derepresija, a ovakav vid kontrole, zasnovan na konformacionoj promjeni represora izazvanoj vezivanjem šećera, je alosterična kontrola. Kao rezultat derepresije, ćelija počinje da sintetiše enzime neophodne za razgradnju laktoze. Kada se razgradi sva prisutna laktoza, a u ćeliju iz okoline ne dospijeva nova, represor se ponovo vezuje za operator i sprečava transkripciju strukturnih gena lac operona. Prema tome, bakterija sintetiše enzime za razgradnju laktoze samo onda kada je laktoza (odnosno alolaktoza) prisutna u ćeliji, a ne i onda kada je nema.
Do sada su poznati mnogi slični primjeri kontrole transkripcije. U svim ovakvim slučajevima, specifičan ligand (kao alolaktoza), koji se naziva induktor, biva prepoznat od strane represora koji imaju visok afinitet prema operatorima. Vezivanje induktora za represor “uključuje” transkripcijsku jedinicu u blizini operatora, slabljenjem veze između represora i operatora. Međutim, poznati su i primjeri kada vezivanje induktora za represor dovodi do takve alosterske promjene represora usljed koje se njegov afinitet prema operatoru povećava, a transkripcijska jedinica ostaje “zaključana”. Znači, u nekim slučajevima, kao što je lac operon, povećana koncentracija induktora stimulira, dok u drugim slučajevima inhibira transkripciju gena. lpak, u oba slučaja radi se o istom regulatornom mehanizmu, jer se u oba slučaja transkripcija odvija u odsustvu regulatornog proteina (represora), a ne odvija u njegovom prisustvu. Zato se ovakav tip kontrole naziva negativna kontrola.
Regulacija transkripcije laktoznog operona - pozitivna kontrola
[uredi | uredi izvor]Alternativa negativnoj regulaciji je pozitivna kontrola, tj. regulacija transkripcije aktivatorima. Neke transkripcione jedinice u genomu E. coli imaju relativno slaba mjesta za vezivanje RNK polimeraze (tzv. slabe promotore), pa je vezivanje ovog enzima moguće samo uz pomoć proteina aktivatora koji se vezuju za DNK u neposrednoj blizini RNK polimeraze.
“Jačina”, tj. efikasnost promotora zavisi od redoslijeda nukleotida u karakterističnim, evolutivno očuvanim dijelovima promotorskog regiona od kojih se jedan nalazi 10, a drugi 35 nukleotida uzvodno od mjesta na kome počinje transkripcija. Najefikasniji bakterijski promotori na poziciji -10 sadrže niz TATAAT, a na -35 niz TTGACA. Molekula RNK polimeraze se specifično vezuje za niz na poziciji -35, dok onaj na poziciji -10 omogućuje da transkripcija započne na nukleotidu +1. Aktivatori se vezuju za nizove nukleotida koji se nalaze u neposrednoj blizini mjesta za koje se vezuje RNK polimeraza i za razliku od represora, koji onemogućuju vezivanje enzima, aktivatori to vezivanje olakšavaju. Po svemu ostalom, oni veoma liče na represore. Oni se često vezuju za induktore koji povećavaju ili smanjuju njihov afinitet prema DNK i na taj način “uključuju” ili “isključuju” odgovarajuće transkripcione jedinice. Ovakav način regulacije aktivnosti gena označen je kao pozitivna kontrola, jer se transkripcija odvija samo u prisustvu regulatornog proteina.
Osnovni izvor energije za bakteriju E. coli je glukoza. Kada je u ćeliji prisutna dovoljna količina glukoze, tada je inhibirana ekspresija svih gena koji kodiraju enzime uključene u razgradnju drugih katabolita, kao što su laktoza, arabinoza i galaktoza. Ovaj fenomen je poznat kao represija katabolita, i njime se sprečava nepotrebno dupliranje enzimskih sistema koji učestvuju u produkciji energije. Otkrivanju ovog fenomena prethodilo je zapažanje da je u prisustvu glukoze u bakterijskoj ćeliji koncentracija cikličnog AMP-a (cAMP-a) manja nego u odsustvu glukoze i da se dodavanjem cAMP-a može prevazići represija katabolita izazvana glukozom. Naime, transport glukoze kroz membranu bakterijske ćelije dovodi do defosforilacije jednog enzima koji mora biti fosforilisan da bi aktivirao adenilat ciklazu, tako da unos glukoze u ćeliju uvek dovodi do snižavanja koncentracije cAMP-a.
Nađeno je da bakterija sadrži protein označen kao CAP (od eng. catabolite gene activator protein) ili CRP (od eng. cAMP receptor protein), koji se sastoji od dvije subjedinice i vezuje cAMP. Vezivanje cAMP-a izaziva veliku konformacionu promjenu ovog proteina, a njegova funkcija je da se u kompleksu s cAMP-om vezuje za lac operon (i neke druge operone), stimulišući njegovu transkripciju u odsustvu lac represora. Prema tome, CAP je pozitivan regulator, odnosno aktivator transkripcije lac operona, za razliku od lac represora koji je negativan regulator. Za sada još uvijek nije sasvim jasno na koji način kompleks CAP-cAMP stimuliše inicijaciju transkripcije na slabom promotoru laktoznog i drugih operona. Jedna mogućnost je direktna interakcija ovog kompleksa s RNK polimerazom, a druga je promjena konformacije DNK u oblasti promotora, koja olakšava inicijaciju transkripcije. U svakom slučaju, ovakav mehanizam regulacije transkripcije lac operona omogućava bakterijskoj ćeliji da u odsustvu glukoze dođe do potrebne energije razgradnjom laktoze, jer kada je koncentracija glukoze u ćeliji niska, koncentracija cAMP-a pa prema tome i kompleksa CAP-cAMP uvijek raste, što dovodi do ubrzane transkripcije lac operona.
Neki regulatorni proteini kod bakterija mogu se vezati na više mjesta u genomu i izvršiti represiju transkripcije jednog operona, a aktivaciju transkripcije drugog, što praktično znači da jedan isti protein može imati i ulogu represora i ulogu aktivatora. Način djelovanja regulatornog proteina zavisi od rastojanja između mjesta na DNK za koje se vezuje regulatorni protein i mjesta za koje se vezuje RNK polimeraza. Ako se ta dva mjesta preklapaju, onda vezivanje regulatornog proteina onemogućuje vezivanje enzima pa protein djeluje kao represor transkripcije. Ukoliko se vezivna mjesta nalaze u blizini, onda vezivanje regulatornog proteina može olakšati vezivanje RNK polimeraze i stimulisati inicijaciju transkripcije pa u tom slučaju protein deluje kao aktivator transkripcije.
Interesantan primjer regulacije transkripcije, u kome jedan isti protein vrši i pozitivnu i negativnu kontrolu u istom operonu, je regulacija transkripcije arabinoznog operona (araBAD operona) E. coli, koji kodira enzime za razgradnju L-arabinoze. Transkripciju ovog operona reguliše kompleks CAP-scAMP i protein AraC (produkt gena araC) koji vezuje L-arabinozu. Mehanizam regulacije transkripcije araBAD operona sastoji se u sljedećem: 1. u odsustvu proteina AraC, RNK polimeraza započinje transkripciju gena araC, što ima za posljedicu sintezu proteina AmC. Operon araBAD se nalazi nizvodno od gena araC i njegova transkripcija se tada održava na bazalnom nivou; 2. kada je koncentracija glukoze u bakterijskoj ćeliji visoka, tada je koncentracija cAMP-a niska, pa je i koncentracija kompleksa CAP-cAMP, također, niska. Protein AmC tada sprečava transkripciju araBAD operona (bakterija nema potrebe da razgrađuje arabinozu, jer dovoljno energije dobija razgradnjom glukoze). Situacija je slična i kada u bakterijskoj ćeliji nema L-arabinoze, jer bakterija nema potrebu za enzimima za razgradnju ovog šećera. Protein AraC se vezuje za tri mjesta. To su araI1 (nalazi se u okviru promotora araBAD operona), araO1 (nalazi se u okviru promotora gena araC) i araO2 (nalazi se u 5' nekodirajućem regionu gena araC). Niz araO1 je operator gena araC i kada se protein AraC za njega veže, transkripcija gena araC je blokirana. Da bi transkripcija operona araBAD bila onemogućena, protein AraC mora biti istovremeno vezan i za segment araI1 i za araO2. Pošto je niz araO2 veoma udaljen od araI1, vjerovatno je da se DNK savija u petlju tako da protein AraC kao dimer ili oligomer može da bude istovremeno vezan za oba ova mjesta, blokirajući transkripciju araBAD operona (negativna regulacija); 3. kada je u ćeliji prisutna L-arabinoza, ona se kao induktor vezuje za protein AraC, dovodeći do takve konformacione promjene ovog proteina da se on radije vezuje za niz araI2 u promotorskom regionu operona araBAD, nego za araO2. To dovodi do otvaranja petlje DNK. Ako je istovremeno koncentracija cAMP-a u ćeliji visoka (u odsustvu glukoze), kompleks CAP-cAMP se vezuje za mjesto između araO1 i araI1. Kombinovano dejstvo dva kompleksa (CAP-cAMP i AraC-arabinoza) uključuje njihovu direktnu interakciju i praćeno je aktivacijom RNK polimeraze, koja počinje da transkribuje araBAD operon (pozitivna kontrola). Gen araC ostaje blokiran proteinom AraC, koji ostaje vezan za araO1.
Regulacija transkripcije triptofanskog operona – atenuacija
[uredi | uredi izvor]Atenuacija je mehanizam kontrole transkripcije kojim bakterija reguliše ekspresiju mnogih operona koji kodiraju enzime uključene u biosintezu aminokiselina. Ovaj mehanizam je bio otkriven kada je izučavana regulacija transkripcije triptofanskog operona (trp operon) bakterije E. coli koji kodira pet polipeptida. Ovih pet polipeptida grade tri enzima koji učestvuju u biosintezi triptofana. Ustanovljeno je da se geni L-P operona koordinisano eksprimiraju pod kontrolom trp represora, dimernog proteina koji se sastoji od dvije identične subjedinice. Trp' represor je produkt gena trpR, koji se nalazi u okviru zasebne transkripcione jedinice. On vezuje L-triptofan gradeći kompleks koji se specifično vezuje za trp operator (trpO) i time 70 puta smanjuje brzinu transkripcije trp operona. Vezivanje triptofana za represor dovodi do takve alosterične promjene represora koja omogućuje njegovo istovremeno vezivanje i za operator. Uz to i sam triptofan formira H-vezu s fosfatnom grupom na DNK, tako da učvršćuje vezu između represora i operatora, odnosno djeluje kao korepresor (sprečava sopstvenu sintezu). Trp represor kontroliše transkripciju ne samo trp operona, već još najmanje dva operona: trpR operona i aroH operona (kodira sintezu jednog od tri izoenzima, koji katalizuju biosintezu horizmata, prekursora aromatičnih aminokiselina).
U početku se smatralo da je sinteza triptofana kod E. coli u potpunosti kontrolisana sistemom trp represor-operator. Međutim, nađeno je da nizvodno od trpO postoji još jedan kontrolni element. Ispred prvog strukturnog gena u trp operonu (trpE) nalazi se čeoni niz (trpL) dužine 162 bp i u okviru njega niz dužine oko 30 bp koji ,također, ima ulogu kontrolnog elementa. U odsustvu triptofana, sintetiše se policistronska iRNK dužine 6720 nukleotida koja obuhvata i čeoni niz trpL. Kada koncentracija triptofana u bakteriji raste, brzina transkripcije trp operona opada kao rezultat stvaranja kompleksa trp represor-korepresor. Među iRNK koje se sintetišu zapaža se povećana proporcija kratkih transkripata dužine oko 140 nukleotida koji odgovaraju 5' kraju trpL niza. To znači da u prisustvu triptofana dolazi do prerane terminacije transkripcije trp operona i zato je kontrolni element odgovoran za ovu pojavu nazvan atenuatorom. Pokazalo se da atenuator sadrži četiri komplementa RNK segmenta koji mogu da formiraju dvije različite intramolekulske zavojnice: između segmenata 2 i 3, odnosno 3 i 4, ali tako da formiranje jedne od njih onemogućava formiranje druge. Segmenti 3 i 4 su bogati nukleotidima G i C, a zajedno s nekoliko uzastopnih U koji slijede, čine normalan terminacioni signal za RNK polimerazu.
Transkripcija se može nastaviti poslije ovog terminacionog signala samo u odsustvu triptofana. Segment I je karakterističan po tome što sadrži dva uzastopna kodona za triptofan (tandemski kodon). Da bi se razumio mehanizam atenuacije trp operona, važno je imati na umu da translacija kod prokariota započinje ubrzo pošto se sintetiše 5'-kraj iRNK i teče uporedo s transkripcijom. Mehanizam atenuacije se sastoji u tome da RNK polimeraza koja je izbjegla represiju započinje transkripciju. Ubrzo poslije transkribovanja Šajn-Dalgarnovog niza (mjesto vezivanja male ribosomske subjedinice) koji se nalazi u okviru čeonog niza trpL, ribosom se vezuje za ovaj niz i započinje translaciju. Ako je u ćeliji prisutan triptofan, tada su prisutni i kompleksi Trp-tRNKTrp, tako da iza RNK polimeraze napreduje i ribosom preko triptofanskog tandemskog kodona u segmentu 1 atenuatora. Ribosom sprečava da se stvori ukosnica izmedu segmenata 1 i 2, ali je zato favorizovano stvaranje ukosnice između segmenata 3 i 4 koji grade terminacioni signal. Transkripcija se zato prerano završava. Ako u ćeliji nema triptofana, ribosom se zaustavlja na triptofanskom tandemskom kodonu u segmentu 1 atenuatora, očekujući da se Trp-tRNKTrp, koje nema, veže za A mjesto. Pošto je ribosom zaostao, omogućeno je stvaranje ukosnice između segmenata 2 i 3, a time je spriječeno stvaranje terminacionog signala, tj. ukosnice između segmenata 3 i 4. RNK polimeraza tada nastavlja transkripciju duž cijelog operona, sintetišući policistronsku iRNK koja će poslužiti kao informacija za sintezu enzima neophodnih za biosintezu triptofana. Ovaj regulatorni mehanizam se zasniva na koncentraciji Trp-tRNKTrp u ćeliji, koja zavisi od koncentracije triptofana. Odsustvo triptofana utječe na ekspresiju trp operona na dva načina: uklanjanjem represora, čime se brzina transkripcije povećava oko 70 puta i atenuacijom, čime se brzina transkripcije povećava još 8-10 puta, tako da transkripcija može biti ukupno ubrzana do 700 puta. Na sličan način je kod bakterija regulisana i transkripcija nekih drugih operona, kao što su his operon (kodira enzime koji učestvuju u biosintezi histidina) i ilv operon (kodira enzime koji učestvuju u biosintezi izoleucina, leucina i valina).[1][6]
Reference
[uredi | uredi izvor]- ^ a b Bajrović K, Jevrić-Čaušević A., Hadžiselimović R., Ed. (2005): Uvod u genetičko inženjerstvo i biotehnologiju. Institut za genetičko inženjerstvo i biotehnologiju (INGEB), Sarajevo, ISBN 9958-9344-1-8.
- ^ Sofradžija A., Šoljan D., Hadžiselimović R. (2004): Biologija 1, Svjetlost, Sarajevo, ISBN 9958-10-686-8.
- ^ Ibrulj S., Haverić S., Haverić A. (2008): Citogenetičke metode – Primjena u medicini . Institut za genetičko inženjerstvo i biotehnologiju (INGEB), Sarajevo,ISBN 978-9958-9344-5-2.
- ^ Berberović LJ., Hadžiselimović R. (1986): Rječnik genetike. Svjetlost, Sarajevo, ISBN 86-01-00723-6.
- ^ Hadžiselimović R. (2005): Bioantropologija – Biodiverzitet recentnog čovjeka. Institut za genetičko inženjerstvo i biotehnologiju (INGEB), Sarajevo, ISBN 9958-9344-2-6.
- ^ a b Kapur Pojskić L., Ed. (2014): Uvod u genetičko inženjerstvo i biotehnologiju, 2. izdanje. Institut za genetičko inženjerstvo i biotehnologiju (INGEB), Sarajevo, ISBN 978-9958-9344-8-3.